


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
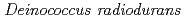
大庭 寛史; 佐藤 勝也; Sghaier, H.; 柳沢 忠*; 鳴海 一成
Extremophiles, 13(3), p.471 - 479, 2009/05
被引用回数:20 パーセンタイル:39.13(Biochemistry & Molecular Biology)デイノコッカス・ラジオデュランスは、PprIが中心的な役割を果たすDNA損傷応答機構を持っており、RecAやPprAの発現を誘導している。このDNA損傷応答機構の特質をさらに明らかにするために、PprI依存性情報伝達経路の新たな因子の同定を試みた結果、コールドショックタンパク質(Csp)と相同性を持つ新たな制御タンパク質を発見し、このタンパク質をPprMと命名した。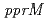 遺伝子破壊株は、
遺伝子破壊株は、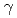 線に著しく感受性を示した。PprMタンパク質は、デイノコッカス・ジオサーマリスやサーマス・サーモフィルスのCSPとともに亜群を形成する独特な分岐群に属していた。また、PprMタンパク質の働きによって、PprAの発現は誘導されるが、RecAの発現は誘導されなかった。PprMタンパク質は、大腸菌のCspDの場合と同様に、生理的条件下で2量体を形成していた。
線に著しく感受性を示した。PprMタンパク質は、デイノコッカス・ジオサーマリスやサーマス・サーモフィルスのCSPとともに亜群を形成する独特な分岐群に属していた。また、PprMタンパク質の働きによって、PprAの発現は誘導されるが、RecAの発現は誘導されなかった。PprMタンパク質は、大腸菌のCspDの場合と同様に、生理的条件下で2量体を形成していた。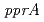 二重遺伝子破壊株は、どちらか一方の遺伝子が欠損した菌株よりも高い放射線感受性を示したことから、PprMタンパク質はPprAタンパク質以外の放射線耐性に重要な未知タンパク質を制御していると考えられた。
二重遺伝子破壊株は、どちらか一方の遺伝子が欠損した菌株よりも高い放射線感受性を示したことから、PprMタンパク質はPprAタンパク質以外の放射線耐性に重要な未知タンパク質を制御していると考えられた。
大庭 寛史; 佐藤 勝也; 菊地 正博; Sghaier, H.; 柳沢 忠*; 鳴海 一成
JAEA-Review 2008-055, JAEA Takasaki Annual Report 2007, P. 57, 2008/11
放射線抵抗性細菌の放射線応答にかかわる新規調節因子として、PprIタンパク質が同定されている。PprIタンパク質はDNA修復促進タンパク質PprAの発現誘導にかかわっているが、PprIタンパク質が膜タンパク質であることから、PprAタンパク質の発現誘導を直接制御している訳ではないことが示唆された。そこで本研究では、PprIタンパク質による直接的制御を受けて、PprAタンパク質の発現を誘導する新規タンパク質を同定することとした。野生株と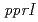 遺伝子破壊株のタンパク質プロファイル解析によって、
遺伝子破壊株のタンパク質プロファイル解析によって、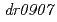 遺伝子が同定された。遺伝子破壊株は線感受性であり、PprAタンパク質の発現誘導の制御が脱抑制されていた。われわれは、PprAタンパク質のモジュレーターの意から、この遺伝子産物をPprMタンパク質と命名した。本研究は、PprMタンパク質がPprAタンパク質の発現誘導制御機構に重要な役割を果たしており、放射線抵抗性細菌のPprIタンパク質に依存した独特な放射線応答機構に関与していることを強く示唆している。
遺伝子が同定された。遺伝子破壊株は線感受性であり、PprAタンパク質の発現誘導の制御が脱抑制されていた。われわれは、PprAタンパク質のモジュレーターの意から、この遺伝子産物をPprMタンパク質と命名した。本研究は、PprMタンパク質がPprAタンパク質の発現誘導制御機構に重要な役割を果たしており、放射線抵抗性細菌のPprIタンパク質に依存した独特な放射線応答機構に関与していることを強く示唆している。
遺伝子の放射線応答プロモーター大庭 寛史; 佐藤 勝也; 柳沢 忠*; 鳴海 一成
JAEA-Review 2006-042, JAEA Takasaki Annual Report 2005, P. 70, 2007/02
放射線抵抗性細菌の優れたDNA2本鎖切断修復には、DNA損傷が生じた後に合成されるタンパク質が必要である。このことは、放射線抵抗性細菌には放射線誘導性のDNA修復機構があることを示している。しかしながら、放射線抵抗性細菌の放射線応答の分子機構を明らかにすることが重要であるにもかかわらず、放射線誘導性タンパク質遺伝子の発現に必要な放射線応答プロモーターについてはほとんど研究されていない。本研究では、放射線で発現が誘導される放射線抵抗性細菌デイノコッカス・ラジオデュランスの遺伝子のプロモーターの放射線応答機構について、ルシフェラーゼ・レポーターアッセイを用いた解析を行った。その結果、放射線応答に必要最小プロモーター領域を同定することに成功した。本研究で同定した遺伝子の放射線応答プロモーターは、今後デイノコッカス・ラジオデュランスの放射線応答機構を全容解明していくうえで非常に役立つものと考えられる。
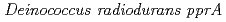 gene
gene大庭 寛史*; 佐藤 勝也*; 柳沢 忠*; 鳴海 一成
Gene, 363, p.133 - 141, 2005/12
被引用回数:37 パーセンタイル:56.73(Genetics & Heredity)デイノコッカス・ラジオデュランス遺伝子の転写開始点が、翻訳開始点上流-156, -154, -22であることを同定した。全ての転写産物の量が放射線照射で増加したことから、少なくとも2つの放射線応答性プロモーターの存在が示唆された。ルシフェラーゼレポーターアッセイにより、遠位プロモーターは翻訳開始点上流-208から-156の間に、近位プロモーターは-57から-22の位置にあることが明らかとなった。-57から-38の領域は、近位プロモーター活性に必須であり、また、-33の位置のチミンが放射線応答に重要な塩基であることがわかった。さらに、遺伝子産物による遺伝子の発現制御がプロモーターレベルで起こることが示唆された。
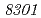 in the radioresistant bacterium
in the radioresistant bacterium Islam, M. S.*; Hua, Y.*; 大庭 寛史; 佐藤 勝也; 菊地 正博; 柳沢 忠*; 鳴海 一成
Genes and Genetic Systems, 78(5), p.319 - 327, 2003/10
被引用回数:16 パーセンタイル:30.57(Biochemistry & Molecular Biology)放射線抵抗性細菌デイノコッカス・ラジオデュランスKD8301株から単離された挿入配列ISを解析した。ISは、IS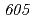 /IS
/IS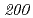 グループに属する新規挿入配列であり、長さが1,736塩基対であった。Inverse PCR法での解析により、ISの挿入標的部位は"TTGAT"であることがわかった。ISラジオデュランスの野生株R
グループに属する新規挿入配列であり、長さが1,736塩基対であった。Inverse PCR法での解析により、ISの挿入標的部位は"TTGAT"であることがわかった。ISラジオデュランスの野生株R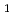 , MR, KRでのISのゲノム内分布を調べたところ、R株の公開ゲノム配列情報と矛盾しており、今回調べたR株ではISが1コピーしか存在しなかった。ISは挿入部位に存在する遺伝子を破壊しており、ゲノムに変異を起こす原因となりうるが、大規模のゲノム再構成を引き起こしてはいなかった。
, MR, KRでのISのゲノム内分布を調べたところ、R株の公開ゲノム配列情報と矛盾しており、今回調べたR株ではISが1コピーしか存在しなかった。ISは挿入部位に存在する遺伝子を破壊しており、ゲノムに変異を起こす原因となりうるが、大規模のゲノム再構成を引き起こしてはいなかった。
佐藤 勝也; 鳴海 一成; 菊地 正博; 北山 滋; 柳沢 忠*; 山本 和生; 渡辺 宏
Journal of Biochemistry, 131(1), p.121 - 129, 2002/01
被引用回数:26 パーセンタイル:39.17(Biochemistry & Molecular Biology)これまでデイノコッカス・ラジオデュランスRecAは,大腸菌とは異なり特別な機能を有しているのではないかと考えられてきた。本論文ではラジオデュランスの野生型及び2種類の変異型RecAを解析し,ラジオデュランスRecAが大腸菌RecAと同等の機能を有していること、当該菌の放射線抵抗性にはRecAによるDNA組換え活性よりも、co-protease活性の方がより重要な役割を担っていることを示した。
is not involved in RecA induction following irradiation鳴海 一成; 佐藤 勝也; 菊地 正博; 舟山 知夫; 柳沢 忠*; 小林 泰彦; 渡辺 宏; 山本 和生
Journal of Bacteriology, 183(23), p.6951 - 6956, 2001/12
被引用回数:91 パーセンタイル:83.56(Microbiology)放射線抵抗性細菌デイノコッカス・ラジオデュランスにおけるLexA蛋白質のRecA誘導への関与について研究を行った。野生株と同様に、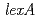 遺伝子破壊株においても線照射後にRecA蛋白質合成の増加が認められたことから、デイノコッカス・ラジオデュランスではLexAがRecAの誘導に関与していないことが示された。
遺伝子破壊株においても線照射後にRecA蛋白質合成の増加が認められたことから、デイノコッカス・ラジオデュランスではLexAがRecAの誘導に関与していないことが示された。
鳴海 一成; 佐藤 勝也; 菊地 正博; 舟山 知夫; 北山 滋; 柳沢 忠*; 渡辺 宏; 山本 和生
Mutation Research; DNA Repair, 435(3), p.233 - 243, 1999/00
放射線抵抗性細菌デイノコッカス・ラジオデュランスのDNA修復能欠損株であるrec30株は放射線に極めて感受性を示し、重要なDNA修復遺伝子に変異が生じているものと考えられる。この変異株のrecA遺伝子座のDNA塩基配列を野生株のものと較べて解析した結果、変異株ではrecA構造遺伝子内に変異があることを突き止め、その正確な変異部位を同定した。また、ラジオデュランスの遺伝子を大腸菌で発現させることは今まで困難であったが、正常及び変異recA遺伝子を大腸菌で大量に発現させることに成功した。さらに、ラジオジュランスの遺伝子を発現している大腸菌の線耐性を測定した結果、ラジオデュランスの正常recA遺伝子が大腸菌のrecA遺伝子の働きを完全に捕らえること、異常recA遺伝子が組換え修復能を完全に失っていることを明らかにした。
; A Novel key protein "PprM"大庭 寛史; Sghaier, H.; 佐藤 勝也; 柳沢 忠*; 鳴海 一成
no journal, ,
デイノコッカス・ラジオデュランスの放射線応答メカニズムの解明が重要であるにもかかわらず、放射線応答メカニズムの上位制御因子であるPprIタンパク質の詳細な機能についてはほとんどわかっていない。そこで、prIタンパク質の機能解析を試みた。ウエスタンブロットとゲルシフトアッセイによりPprIは直接的にや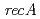 の発現を制御していないことが示唆された。そして、2D-PAGEによりPprMタンパク質を新たに見つけた。遺伝子破壊株は線に対して高感受性であることがわかった。また、ウエスタンブロットによってPprMはRecAとPprAの発現を制御していることが明らかになり、さらに、の発現をプロモーターレベルで制御していることを明らかにした。これらの結果はPprMはデイノコッカス・ラジオデュランスのPprIによって制御されている放射線応答メカニズムに関与しているといえる。
の発現を制御していないことが示唆された。そして、2D-PAGEによりPprMタンパク質を新たに見つけた。遺伝子破壊株は線に対して高感受性であることがわかった。また、ウエスタンブロットによってPprMはRecAとPprAの発現を制御していることが明らかになり、さらに、の発現をプロモーターレベルで制御していることを明らかにした。これらの結果はPprMはデイノコッカス・ラジオデュランスのPprIによって制御されている放射線応答メカニズムに関与しているといえる。
遺伝子の放射線応答プロモーターの解析大庭 寛史*; 佐藤 勝也*; 柳沢 忠*; 鳴海 一成
no journal, ,
放射線抵抗性細菌デイノコッカス・ラジオデュランスの優れたDNA二本鎖切断修復には、DNA損傷が生じた後に合成されるタンパク質が必要である。これまでに、デイノコッカス・ラジオデュランスの放射線抵抗性に中心的役割を担う新規放射線誘導タンパク質PprAを同定した。しかし、放射線応答の分子機構を明らかにすることが重要であるにもかかわらず、放射線誘導性タンパク質の放射線応答プロモーターについてはほとんど明らかにされていない。そこで、遺伝子プロモーターの放射線応答機構について解析を行った。ルシフェラーゼ遺伝子を用いたレポーターアッセイにより、放射線照射による遺伝子プロモーター活性の変動を解析したところ、遠位のプロモーターは-208から-156領域に、近位のプロモーターは-57から-22領域に存在すること、そして、-57から-38領域が最小必要プロモーター領域であることを明らかにした。本研究で同定した遺伝子の放射線応答プロモーターは、今後デイノコッカス・ラジオデュランスの放射線応答機構を全容解明していくうえで非常に役立つと考えられた。
遺伝子の放射線応答プロモーターの解析大庭 寛史*; 佐藤 勝也*; 柳沢 忠*; 鳴海 一成
no journal, ,
放射線抵抗性細菌デイノコッカス・ラジオデュランスの優れたDNA二本鎖切断修復には、DNA損傷が生じた後に合成されるタンパク質が必要である。これまでに、デイノコッカス・ラジオデュランスの放射線抵抗性に中心的役割を担う新規放射線誘導性タンパク質PprAを同定した。しかしながら、放射線応答の分子機構を明らかにすることが重要であるにもかかわらず、放射線誘導性タンパク質の放射線応答プロモーターについてほとんど明らかにされていない。そこで本研究では、遺伝子プロモーターの放射線応答機構について解析を行った。放射線応答に関与するプロモーター領域を同定するため、ルシフェラーゼ遺伝子を用いたレポーターアッセイにより、放射線照射による遺伝子プロモーター活性変動を解析した。遠位のプロモーターは-208から-156領域に、近位のプロモーターは-57から-22領域に存在すること、そして、-57から-38領域が最小必要プロモーター領域であることを明らかにした。さらに、部位特異的変異導入法による解析から、-33位のチミンが放射線応答プロモーターの主要な塩基であることを明らかにした。また、遺伝子産物は放射線応答において上位制御因子と考えられているが、遺伝子破壊株における遺伝子プロモーター活性を解析したところ、遺伝子発現は遺伝子産物によりプロモーターレベルで制御されていることが明らかになった。
遺伝子の放射線応答プロモーターの解析大庭 寛史; 佐藤 勝也; 柳沢 忠*; 鳴海 一成
no journal, ,
放射線抵抗性細菌の優れたDNA2本鎖切断修復には、DNA損傷が生じた後に合成されるタンパク質が必要である。放射線抵抗性細菌の放射線応答の分子機構を明らかにすることが重要であるにもかかわらず、放射線誘導性タンパク質の放射線応答プロモーターについてはほとんど明らかにされていない。本研究では、放射線で発現が誘導されるデイノコッカス・ラジオデュランス遺伝子のプロモーターの放射線応答機構について、ルシフェラーゼ・レポーターアッセイを用いた解析を行った。その結果、放射線応答に必要最小プロモーター領域を同定することに成功した。
大庭 寛史; 佐藤 勝也; 菊地 正博; 鳴海 一成; Sghaier, H.*; 柳沢 忠*
no journal, ,
放射線抵抗性細菌の放射線応答にかかわる新規調節因子として、PprIタンパク質が同定されている。PprIタンパク質はDNA修復促進タンパク質PprAの発現誘導にかかわっているが、PprIタンパク質が膜タンパク質であることから、PprAタンパク質の発現誘導を直接制御している訳ではないことが示唆された。そこで本研究では、PprIタンパク質による直接的制御を受けて、PprAタンパク質の発現を誘導する新規タンパク質を同定することとした。野生株と遺伝子破壊株のタンパク質プロファイル解析によって、遺伝子が同定された。遺伝子破壊株は線感受性であり、PprAタンパク質の発現誘導の制御が脱抑制されていた。われわれは、PprAタンパク質のモジュレーターの意から、この遺伝子産物をPprMタンパク質と命名した。本研究は、PprMタンパク質がPprAタンパク質の発現誘導制御機構に重要な役割を果たしており、放射線抵抗性細菌のPprIタンパク質に依存した独特な放射線応答機構に関与していることを強く示唆している。
