


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
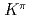 = 8
= 8 two-quasineutron isomer in
two-quasineutron isomer in 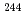 Pu
PuHota, S.*; Tandel, S.*; Chowdhury, P.*; Ahmad, I.*; Carpenter, M. P.*; Chiara, C. J.*; Greene, J. P.*; Hoffman, C. R.*; Jackson, E. G.*; Janssens, R. V. F.*; et al.
Physical Review C, 94(2), p.021303_1 - 021303_5, 2016/08
被引用回数:9 パーセンタイル:54.19(Physics, Nuclear)Puにおける = 8アイソマーからの崩壊と集団的バンド構造が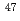 Tiと
Tiと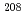 Pbのビームによる深部非弾性散乱実験によって調べられた。バンド内の正確な
Pbのビームによる深部非弾性散乱実験によって調べられた。バンド内の正確な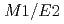 分岐比の測定によって、偶
分岐比の測定によって、偶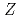 ,
, 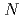 =150アイソトーンにおける = 8二準中性子アイソマーが9/2
=150アイソトーンにおける = 8二準中性子アイソマーが9/2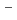 [734]
[734]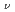
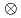 7/2
7/2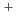 [624]の配位であることを確かめた。=152における変形シェルギャップ近傍のこれらのアイソマーは、超重核の一粒子エネルギーの理論的な予言において重要なベンチマークとなる。
[624]の配位であることを確かめた。=152における変形シェルギャップ近傍のこれらのアイソマーは、超重核の一粒子エネルギーの理論的な予言において重要なベンチマークとなる。
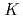 isomer in
isomer in 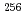 Rf
RfRobinson, A. P.*; Khoo, T. L.*; Seweryniak, D.*; Ahmad, I.*; 浅井 雅人; Back, B. B.*; Carpenter, M. P.*; Chowdhury, P.*; Davids, C. N.*; Greene, J.*; et al.
Physical Review C, 83(6), p.064311_1 - 064311_7, 2011/06
被引用回数:35 パーセンタイル:85.00(Physics, Nuclear)104番元素Rfの励起状態に半減期17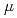 秒の核異性体があることを実験的に明らかにした。実験はアルゴンヌ国立研究所の反跳核分離装置を用いて行い、検出器に打ち込まれたRfと同時計数する内部転換電子を測定することで同定した。核異性体の生成率から、この核異性体は中性子数152の原子核で通常観測される2準粒子状態の核異性体ではなく、4準粒子状態の核異性体であると考えられる。2準粒子状態の核異性体が観測されなかった理由は、その核異性体が核分裂によって崩壊し、その半減期がRfの基底状態の半減期に近いと考えれば説明できる。あるいは、原子核の4重極変形度が104番元素から突然小さくなり、2準粒子状態の核異性体がまったく存在しない、という可能性も考えられる。
秒の核異性体があることを実験的に明らかにした。実験はアルゴンヌ国立研究所の反跳核分離装置を用いて行い、検出器に打ち込まれたRfと同時計数する内部転換電子を測定することで同定した。核異性体の生成率から、この核異性体は中性子数152の原子核で通常観測される2準粒子状態の核異性体ではなく、4準粒子状態の核異性体であると考えられる。2準粒子状態の核異性体が観測されなかった理由は、その核異性体が核分裂によって崩壊し、その半減期がRfの基底状態の半減期に近いと考えれば説明できる。あるいは、原子核の4重極変形度が104番元素から突然小さくなり、2準粒子状態の核異性体がまったく存在しない、という可能性も考えられる。
Seweryniak, D.*; Khoo, T. L.*; Ahmad, I.*; Kondev, F. G.*; Robinson, A.*; Tandel, S. K.*; 浅井 雅人; Back, B. B.*; Carpenter, M. P.*; Chowdhury, P.*; et al.
Nuclear Physics A, 834(1-4), p.357c - 361c, 2010/03
被引用回数:8 パーセンタイル:46.62(Physics, Nuclear)陽子数100,中性子数152近傍の原子核における一粒子軌道エネルギーの実験値は、超重核領域の殻構造を予測する理論計算の検証に重要な役割を果たす。われわれは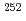 No及び
No及び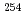 Noに2準粒子状態の高K核異性体を観測し、それらのエネルギーから陽子数100近傍における陽子の一粒子軌道エネルギーを評価することに成功した。また
Noに2準粒子状態の高K核異性体を観測し、それらのエネルギーから陽子数100近傍における陽子の一粒子軌道エネルギーを評価することに成功した。また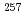 Rfに3準粒子状態の高K核異性体を発見し、Rfの
Rfに3準粒子状態の高K核異性体を発見し、Rfの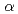 崩壊の実験データから中性子の一粒子軌道エネルギーも評価した。得られた実験値をさまざまな理論計算の予測値と比較した結果、Woods-Saxonポテンシャルを用いた計算が最もよく実験値を再現することを見いだした。
崩壊の実験データから中性子の一粒子軌道エネルギーも評価した。得られた実験値をさまざまな理論計算の予測値と比較した結果、Woods-Saxonポテンシャルを用いた計算が最もよく実験値を再現することを見いだした。
RfQian, J.*; Heinz, A.*; Khoo, T. L.*; Janssens, R. V. F.*; Peterson, D.*; Seweryniak, D.*; Ahmad, I.*; 浅井 雅人; Back, B. B.*; Carpenter, M. P.*; et al.
Physical Review C, 79(6), p.064319_1 - 064319_13, 2009/06
被引用回数:35 パーセンタイル:85.51(Physics, Nuclear)アルゴンヌ国立研究所の反跳核分離装置を用いて、Rfの線,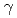 線,内部転換電子測定実験を行った。Rfの励起状態に、高いK量子数を持った3準粒子状態と解釈される半減期160秒の新核異性体を発見した。また、崩壊の抑止係数より、Rfの娘核である
線,内部転換電子測定実験を行った。Rfの励起状態に、高いK量子数を持った3準粒子状態と解釈される半減期160秒の新核異性体を発見した。また、崩壊の抑止係数より、Rfの娘核である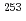 Noの1準粒子状態の中性子軌道配位を同定した。中性子数151核の1/2
Noの1準粒子状態の中性子軌道配位を同定した。中性子数151核の1/2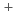 [620]準位の励起エネルギーの系統性より、中性子数152の変形閉殻の大きさが、原子番号が増えるにつれて大きくなることを明らかにした。
[620]準位の励起エネルギーの系統性より、中性子数152の変形閉殻の大きさが、原子番号が増えるにつれて大きくなることを明らかにした。
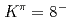 isomers and
isomers and 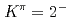 octupole vibrations in
octupole vibrations in 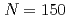 shell-stabilized isotones
shell-stabilized isotonesRobinson, A. P.*; Khoo, T. L.*; Ahmad, I.*; Tandel, S. K.*; Kondev, F. G.*; 中務 孝*; Seweryniak, D.*; 浅井 雅人; Back, B. B.*; Carpenter, M. P.*; et al.
Physical Review C, 78(3), p.034308_1 - 034308_6, 2008/09
被引用回数:53 パーセンタイル:91.13(Physics, Nuclear)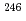 Cm及びNoの励起準位に、量子数を持ち、の八重極振動回転バンドを経由して崩壊する核異性体を発見した。これらの中性子数核における及び2準位は、原子番号
Cm及びNoの励起準位に、量子数を持ち、の八重極振動回転バンドを経由して崩壊する核異性体を発見した。これらの中性子数核における及び2準位は、原子番号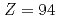
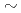 102の範囲においてほぼ一定の励起エネルギーを持つことから、中性子の励起に起因する準位と考えられる。ただしCmの2準位だけは例外的に低い励起エネルギーを持ち、中性子の励起に加えて陽子の配位の影響を受けていると考えられる。
102の範囲においてほぼ一定の励起エネルギーを持つことから、中性子の励起に起因する準位と考えられる。ただしCmの2準位だけは例外的に低い励起エネルギーを持ち、中性子の励起に加えて陽子の配位の影響を受けていると考えられる。
Ahmad, S.*; 河野 秀俊; Ara zo-Bravo, M. J.*; 皿井 明倫*
zo-Bravo, M. J.*; 皿井 明倫*
Nucleic Acids Research, 34(Suppl.2), p.W124 - W127, 2006/07
被引用回数:31 パーセンタイル:48.95(Biochemistry & Molecular Biology)転写因子などのDNA結合蛋白質はDNA配列に特異的に結合する。DNA結合蛋白質のDNA配列の認識は、おもに静電相互作用,水素結合,ファンデルワールス相互作用などの塩基とアミノ酸残基の直接的な相互作用によって行われていると考えられてきた。しかし、多くの蛋白質-DNA複合体構造が明らかになるにつれ、直接的な相互作用の数が少ないこと,直接に蛋白質と相互作用をしていない塩基配列を変えても結合強度が変わることなどが明らかになり、直接的でない認識、つまり、間接認識の重要性も認知されるようになってきた。間接認識とは、配列に依存したDNAの特異構造や曲がりやすさなどの構造情報や物性情報を通してDNA配列を蛋白質が間接的に認識することをさす。したがって、蛋白質とDNAの認識機構を解明するには、この両者の寄与を明らかにする必要がある。しかしながら、直接認識と間接認識の寄与を定量化することが難しいため、その寄与バランスはよくわかっていない。われわれは、両者の寄与を定量化する方法を開発し、研究者がさまざまな蛋白質-DNA複合体についてその寄与バランスを知ることができるように、寄与バランスを計算できるツールをインターネット上に公開した。
Arazo-Bravo, M. J.*; 藤井 聡*; 河野 秀俊; Ahmad, S.*; 皿井 明倫*
Journal of the American Chemical Society, 127(46), p.16074 - 16089, 2005/11
被引用回数:46 パーセンタイル:72.48(Chemistry, Multidisciplinary)すべてのテトラマー配列(136通り)を12merのDNA配列の真中にもつDNAの分子動力学計算を行い、DNA構造,ダイナミクスの配列依存性を解析した。その結果、ピリミジン-プリンステップを持つ配列は非常に前後の配列の影響を受けやすいことがわかった。また、分子動力学計算から得られた構造アンサンブルから統計ポテンシャルを計算し、簡便にDNAの構造エネルギーを評価できるようにした。
河野 秀俊; Kumar, S.*; Ahmad, S.*; Arazo-Bravo, M. J.*; 藤井 聡*; 郷 信広; 皿井 明倫*
no journal, ,
転写因子などのDNA結合蛋白質はDNA配列に特異的に結合する。DNA結合蛋白質のDNA配列の認識は、おもに静電相互作用,水素結合,ファンデルワールス相互作用などの塩基とアミノ酸残基の直接的な相互作用によって行われていると考えられてきた。しかし、多くの蛋白質-DNA複合体構造が明らかになるにつれ、直接的な相互作用の数が少ないこと,直接に蛋白質と相互作用をしていない塩基配列を変えても結合強度が変わることなどが明らかになり、直接的でない認識、つまり、間接認識の重要性も認知されるようになってきた。間接認識とは、配列に依存したDNAの特異構造や曲がりやすさなどの構造情報や物性情報を通してDNA配列を蛋白質が間接的に認識することをさす。したがって、蛋白質とDNAの認識機構を解明するには、この両者の寄与を明らかにする必要がある。しかしながら、直接認識と間接認識の寄与を定量化することが難しいため、その寄与バランスはよくわかっていない。われわれは、両者の寄与を定量化する方法を開発し、さまざまな蛋白質-DNA複合体についてその寄与バランス調べた。直接認識は、蛋白質-DNAの複合体構造から抽出されたアミノ酸と塩基の空間分布から計算されるknowledge-based potentialを、間接認識では、複合体のDNA構造の配列依存性から抽出されるknowledge-based potential及びさまざまなDNA配列の分子動力学計算による構造アンサンブルから計算されるpotential of mean forceを用いて、それぞれの寄与をエネルギーとして定量化した。計算したエネルギーは、ランダムDNA配列とのエネルギーの差としてそれぞれで標準化し(Z-score化)、直接認識と間接認識の寄与を評価した。本発表では、約200の蛋白質-DNA複合体における寄与バランスを報告する。
Ngahu, A.*; 上野 卓哉*; Ahmad, S.*; 河野 秀俊; 皿井 明倫*
no journal, ,
酵母ゲノムにおいて、DNA結合蛋白質を推定し、その推定したDNA結合蛋白質のターゲット配列をさらに推定した。これにより、どの遺伝子がどの遺伝子と関連があるか予測することができる。酵母の場合、約150のDNA結合蛋白質が推定された。同じターゲット配列を持つ遺伝子は、細胞周期の同時期に働くと考えられており、その産物である蛋白質は、お互いに相互作用する可能性が高い。これをもとに、蛋白質-蛋白質相互作用の推定に応用できる。
皿井 明倫*; 上野 卓哉*; Ngahu, A.*; Ahmad, S.*; 河野 秀俊
no journal, ,
500以上の生物種のゲノム解析から膨大な機能未知の遺伝子がもたらされている。これらのデータから転写因子やそのターゲット予測をゲノムスケールで行うことは、遺伝子発現制御のメカニズムを明らかにするうえで重要である。われわれは、機能未知の遺伝子からDNA結合蛋白質や転写因子のターゲットを予測する方法を開発してきた。DNA結合蛋白質の予測では、DNA結合蛋白質の配列データ,組成やコンテクスト,配列アラインメントの進化的情報や、2次構造,露出表面積,電荷分布の特性(全電荷,双極子等)などの構造情報に基づいて予測する方法を開発した。一方、転写因子のターゲット予測では、蛋白質・DNA複合体の構造データを統計的に解析することにより、アミノ酸と塩基の相互作用による直接認識及びDNAのコンフォメーションによる間接認識の統計ポテンシャルを導出し、それらをもとに予測する方法を開発した。このほか、配列情報を用いる方法,蛋白質・DNA結合データを用いる方法,計算機シミュレーションなどを組合せて精度を上げようとしている。これらの方法をゲノムレベルの予測に応用するため、酵母ゲノムについて解析を行った。この解析のため、まず酵母に関する種々の情報を統合したデータベースを構築した。このデータベースの情報をもとにDNA結合蛋白質や転写因子のターゲット予測を行い、実験との比較解析を進めている。
