


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
奥津 賢一*; 山下 琢磨*; 木野 康志*; 宮下 湖南*; 安田 和弘*; 岡 壽崇; 岡田 信二*; 佐藤 元泰*
JJAP Conference Proceedings (Internet), 9, p.011003_1 - 011003_7, 2023/00
ミュオン触媒核融合において、負ミュオンは水素同位体の核融合の触媒として働くことが知られている。本研究では、ミュオン触媒核融合サイクルにおける重水素・三重水素・ミュオンの共鳴状態(dt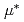 )をルンゲクッタ法による逐次計算により分析した。その結果、dtの生成が核融合反応を促進させることがわかった。
)をルンゲクッタ法による逐次計算により分析した。その結果、dtの生成が核融合反応を促進させることがわかった。
奥津 賢一*; 山下 琢磨*; 木野 康志*; 中島 良太*; 宮下 湖南*; 安田 和弘*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
Fusion Engineering and Design, 170, p.112712_1 - 112712_4, 2021/09
被引用回数:3 パーセンタイル:31.52(Nuclear Science & Technology)水素同位体を利用したミュオン触媒核融合(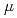 CF)では、核融合によって2.2sの寿命を持つミュオンが再放出され、それが次の標的と新たな核融合を引き起こす。我々は、水素・重水素混合固体から放出されたミュオンを収集して輸送する同軸輸送管を新たに開発し、輸送のための加速電圧などについて検討したので報告する。
CF)では、核融合によって2.2sの寿命を持つミュオンが再放出され、それが次の標的と新たな核融合を引き起こす。我々は、水素・重水素混合固体から放出されたミュオンを収集して輸送する同軸輸送管を新たに開発し、輸送のための加速電圧などについて検討したので報告する。
山下 琢磨*; 奥津 賢一*; 木野 康志*; 中島 良太*; 宮下 湖南*; 安田 和弘*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
Fusion Engineering and Design, 169, p.112580_1 - 112580_5, 2021/08
被引用回数:3 パーセンタイル:31.52(Nuclear Science & Technology)重水素・三重水素混合固体標的に負ミュオン()を入射し、ミュオン触媒核融合反応(CF)の時間発展をルンゲクッタ法によって計算した。核融合によって生成する中性子の強度や、固体標的から真空中に放出されるミュオン量を最大化する三重水素含有率を明らかにした。
安達 基泰; 平山 裕士; 清水 瑠美; 佐藤 勝也; 鳴海 一成*; 黒木 良太
Protein Science, 23(10), p.1349 - 1358, 2014/10
被引用回数:10 パーセンタイル:26.31(Biochemistry & Molecular Biology)DNAの修復に関与する多機能なPprAは、デイノコッカスラジオデュランスの高度な放射線耐性を促進する重要なタンパク質である。PprAによる放射線耐性機構を解明するために、大腸菌で発現した組換え型PprAと2本鎖DNAとの相互作用解析を実施した。ゲルシフトアッセイにより、PprAとスーパーコイル型のpUC19 DNAとの複合体のゲルシフトが2極性であり、それが1mMのMg, Ca, Srイオンで促進されることが示された。シフトしたバンドの相対的な割合からPprAとスーパーコイル型のpUC19 DNAとの複合体形成のヒル係数および解離定数を計算したところ、1mM Mgイオンの存在下で、それぞれ3.3と0.6Mであった。このことは、PprAがスーパーコイル型のpUC19 DNAに少なくとも281分子結合していることを示しており、ゲル濾過での分離後にUV吸収で見積もられた値と一致した。この結果は、PprAが2本鎖DNAに沿って、直鎖状に重合して結合していることを示唆している。一方で、直鎖状の2本鎖DNAとニックがある環状DNAのバンドシフトに関しては、飽和が見られず、1.3M以上のPprA濃度の時、さらに大きな複合体の形成が確認された。この結果は、DNAに結合したPprAが、ダメージを受けたDNAの末端の会合を濃度依存的に促進していることを示している。
阪東 恭子*; 斉藤 健; 佐藤 剛一*; 田中 智章*; Dumeignil, F.*; 今村 元泰*; 松林 信行*; 島田 広道*
Topics in Catalysis, 18(1-2), p.59 - 65, 2002/01
CO水素化反応時におけるRh担持Yゼオライト触媒に関し、XAFS分析法によりその場観察を行った。分析の結果、低温ではRhY触媒は、LiをドーピングしたRhY触媒よりも、より還元されやすいことが明らかとなった。低温でのRhの還元には、Rh酸化物の形成が関与していることが明らかとなり、この現象はLiをドーピングしたRhY触媒では観察されなかった。反応中でのRh粒子の大きさは、RhY触媒では1.3nm,LiをドーピングしたRhY触媒では0.8nmであった。CO反応の場合だけでなく、乾燥空気にさらした場合でも、RhY触媒よりもLiをドーピングしたRhY触媒の方が、より構造変化を受けやすいことも明らかとなった。これらの違いはLiドーピングしたRhY触媒におけるLi酸化物の影響が関与している可能性によるものと結論された。
CF実験におけるミュオン特性X線を用いた放出ミュオンの観測奥津 賢一*; 木野 康志*; 中島 良太*; 宮下 湖南*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
ミュオン触媒核融合(CF)は素粒子の一つである負ミュオンが触媒のように振舞いながら水素同位体間の核融合を起こす反応である。CFは、反応後のミュオンの運動エネルギー分布幅が小さい点や単色の中性子を放出する性質から、非破壊検査に威力を発揮できる運動量の揃った高品質なミュオンビーム源や走査ミュオン顕微鏡などへの応用が期待される。固体水素標的から核反応後に放出される再生ミュオンの運動エネルギー分布の測定法について検討した。
宮下 湖南*; 奥津 賢一*; 木野 康志*; 中島 良太*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
ミュオン触媒核融合によって発生する再生ミュオンの運動エネルギー分布を直接測定することを目指している。放射線輸送計算コードPHITSのシミュレーションに基づいて実験系を設計し、J-PARCにおいてミュオンビーム照射実験を行い、得られた実験結果と計算結果の比較を行ったので報告する。
中島 良太*; 奥津 賢一*; 木野 康志*; 宮下 湖南*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
核反応後に放出される再生ミュオンは、数keVから10keV程度の運動エネルギー分布を持つ。この運動エネルギー分布測定には、重水素膜から放出された再生ミュオンをチタン薄膜まで輸送し、チタン原子から放出される特性ミュオンX線を検出する必要がある。実験系の構築のため、本研究では、イオン軌道計算ソフトウェアSIMIONを用いて、重水素膜からチタン薄膜までの輸送シミュレーションを行った。
宮下 湖南*; 奥津 賢一*; 木野 康志*; 中島 良太*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
ミュオン触媒核融合後に放出される再生ミュオンの運動エネルギー分布測定を行うと、核反応の瞬間のdd (d: 重水素、: ミュオン)の波動関数に関する現象を観察することができる。本研究では、バックグラウンドの原因とその低減方法、シミュレーションコードを用いた輸送する再生ミュオンの軌道などについて議論する。
中島 良太*; 奥津 賢一*; 木野 康志*; 宮下 湖南*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
ミュオン触媒核反応後に放出される再生ミュオンの運動エネルギー分布測定のための実験系を開発している。運動エネルギー分布測定のためには、再生ミュオンを固体水素ターゲットから離れたところまで輸送し、Ti薄膜に衝突させる必要がある。本発表では、電場を用いたミュオン輸送について、荷電粒子軌跡計算ソフトウェアSIMIONと粒子輸送計算コードPHITSで検討した。
奥津 賢一*; 木野 康志*; 中島 良太*; 宮下 湖南*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
電子の207倍の質量を持ち、2.2sの寿命で崩壊するミュオンは、電子より強く原子核同士を結びつけることができるため、2つの水素同位体核とミュオンが形成するミュオン分子内ではミュオン触媒核融合とよばれる核反応が起きる。ミュオン触媒核融合反応後に放出されるミュオン(再生ミュオン)の運動エネルギー分布を測定することで核反応の際のミュオン分子軌道の変化を調べることができる。本講演では、再生ミュオンの検出のために現在開発しているミュオン触媒核融合用水素固体標的系と電場によるミュオン輸送シミュレーションの結果などを報告する。
宮下 湖南*; 奥津 賢一*; 木野 康志*; 中島 良太*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
負ミュオンが重水素分子イオン(D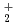 )様のミュオン分子ddを作ると、核間距離はDの1/207程度になり、dd分子内で核融合反応が起きる。我々はこのミュオン触媒核融合(CF)時に放出される再生の運動エネルギー分布の測定を計画しているが、バックグラウンド(BG)が大きいことが問題となっていた。放射線輸送コードPHITSと測定結果の比較から、主要なBGの発生源がサーマルシールドの標的手前側であることがわかったので、BGの発生源を同軸輸送管の軸から離し、X線検出器と発生源の間に遮蔽体を設置できるようサーマルシールドの形状を修正した。その結果、BGを最大で2%未満に抑えることが可能になると見積もられた。
)様のミュオン分子ddを作ると、核間距離はDの1/207程度になり、dd分子内で核融合反応が起きる。我々はこのミュオン触媒核融合(CF)時に放出される再生の運動エネルギー分布の測定を計画しているが、バックグラウンド(BG)が大きいことが問題となっていた。放射線輸送コードPHITSと測定結果の比較から、主要なBGの発生源がサーマルシールドの標的手前側であることがわかったので、BGの発生源を同軸輸送管の軸から離し、X線検出器と発生源の間に遮蔽体を設置できるようサーマルシールドの形状を修正した。その結果、BGを最大で2%未満に抑えることが可能になると見積もられた。
中島 良太*; 奥津 賢一*; 木野 康志*; 宮下 湖南*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
ミュオン触媒核反応後に放出される再生ミュオンの運動エネルギー分布測定のための実験系を開発している。再生ミュオンを同軸輸送管によって制動放射線などのバックグラウンドが低い場所まで輸送するには、再生ミュオンの運動エネルギーや同軸管への入射条件などを最適化する必要がある。荷電粒子軌跡計算ソフトウェアSIMIONを用いて、固体水素標的から放出された再生ミュオンの軌跡を計算し、再生ミュオンの輸送効率や放出位置依存性について議論した。
池本 恵*; 染川 純*; 根木 新太*; 小西 蓮*; 中島 良太*; 奥津 賢一*; 木野 康志*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; et al.
no journal, ,
加速器で生成した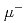 を多段の薄膜で減速し、収束してミュオンビームを高品質化する研究を進めている。本研究では、約4MeVのを厚さ0.5mmのSi薄膜に入射し、数keV程度に減速させたを静電場で引き出した後に、4層の半休型メッシュ電極によって収束させる実験の数値シミュレーションを行った。輸送中のの軌道は薄膜で減速後の運動エネルギーに依存しており、10keVで発射された一様分布の(直径35.2mm)は後段のMg板の上で直径11mm程度に収束するが、15keV以上のエネルギーを持つものは半休型メッシュ電極到着後も収束されるに直線的に運動してMg板に到達することがわかった。メッシュ電極に薄膜を付加することで、さらに集束径を小さく出来ると期待される。この計算に基づいてJ-PARCでの実験を実施している。
を多段の薄膜で減速し、収束してミュオンビームを高品質化する研究を進めている。本研究では、約4MeVのを厚さ0.5mmのSi薄膜に入射し、数keV程度に減速させたを静電場で引き出した後に、4層の半休型メッシュ電極によって収束させる実験の数値シミュレーションを行った。輸送中のの軌道は薄膜で減速後の運動エネルギーに依存しており、10keVで発射された一様分布の(直径35.2mm)は後段のMg板の上で直径11mm程度に収束するが、15keV以上のエネルギーを持つものは半休型メッシュ電極到着後も収束されるに直線的に運動してMg板に到達することがわかった。メッシュ電極に薄膜を付加することで、さらに集束径を小さく出来ると期待される。この計算に基づいてJ-PARCでの実験を実施している。
吉村 直記*; 佐藤 令奈*; 安達 基泰; 黒木 良太; 加藤 悦子*; 城所 俊一*
no journal, ,
低温ショック蛋白質(CSP)は、低温ストレスにより細胞内で発現し、一本鎖核酸と結合することで、核酸の高次構造形成を抑えて、遺伝情報の転写・翻訳への障害を抑制すると考えられている。CSPの研究報告は幾つかあるが、熱力学的に結合を解析した報告は少ない。また、結合の塩基依存性は報告されているが、明らかになった複合体立体構造はすべてオリゴチミジンとの複合体である。本研究では、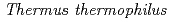 HB8株由来低温ショック蛋白質(以下、
HB8株由来低温ショック蛋白質(以下、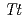 CSP)とTCTTTTTの配列の一本鎖核酸を用いて、滴定型熱量測定法により相互作用をX線構造解析により複合体構造を評価した。
CSP)とTCTTTTTの配列の一本鎖核酸を用いて、滴定型熱量測定法により相互作用をX線構造解析により複合体構造を評価した。
山田 貢; 安達 基泰; 佐藤 勝也; 玉田 太郎; 由良 敬*; 黒木 良太; 鳴海 一成
no journal, ,
PprAは放射線抵抗性細菌デイノコッカスラジオデュランスより単離された放射線誘導性DNA修復促進蛋白質であり、DNAに生じた単鎖切断部位及び二本鎖切断部位を認識して結合し、DNA末端再結合反応を促進する。大腸菌発現系を用いて調製した組換えPprAを精製し、PprAの結晶化を行ったところPprAの結晶化にはタンパク質表面に露出しているリジン残基の化学修飾が有効であることを見いだした。放射光を用いた予備的なX線回折実験を行ったところ、低分解能ながら回折点が確認され、a=b=c=285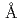 ,
, 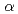 =
=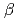 =
=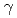 =90
=90 のprimitive cubicであることが明らかになった。またPprA及び化学修飾を施したPprAとDNAとの相互作用をアガロースゲルシフトアッセイとゲル濾過により検討したところ、化学修飾の前後でDNAに対する結合特異性に変化が生じていることを見いだし、その結果をもとに部位特異的変異法を用いて各種変異体PprAを創出し、DNAとの相互作用を解析した。本発表では構造決定に向けた結晶解析の進捗状況並びにDNAとの相互作用実験によって得られた知見を報告する。
のprimitive cubicであることが明らかになった。またPprA及び化学修飾を施したPprAとDNAとの相互作用をアガロースゲルシフトアッセイとゲル濾過により検討したところ、化学修飾の前後でDNAに対する結合特異性に変化が生じていることを見いだし、その結果をもとに部位特異的変異法を用いて各種変異体PprAを創出し、DNAとの相互作用を解析した。本発表では構造決定に向けた結晶解析の進捗状況並びにDNAとの相互作用実験によって得られた知見を報告する。
安達 基泰; 玉田 太郎; 佐藤 勝也; 由良 敬; 鳴海 一成; 黒木 良太
no journal, ,
放射線抵抗性細菌デイノコッカスは、ヒト細胞の約1000倍もの放射線抵抗性を示す。PprAはデイノコッカスより単離された新規なDNA修復促進蛋白質であり、高度放射線抵抗性において最も重要な役割を担っていることが報告されている。本研究は、PprAの構造と機能の関係を解明することを目的としている。大腸菌発現系を用いて調製した組換えPprAを精製し、DNAとの相互作用をアガロースゲルシフトアッセイとゲル濾過により検討した。その結果、PprAは1分子のDNA(pUC19, 2686bp)に少なくとも約280分子の結合が可能であること,PprAとDNAの複合体形成が、低濃度のMg, CaあるいはSrイオンにより促進されること,PprAの多量体構造の形成及びPprAとDNAとの複合体の形成の両方に塩濃度依存性(0-0.4M,酢酸ナトリウム)があることが示された。また、直鎖状の2本鎖DNAとPprAが複合体を形成する場合、PprAの濃度上昇に伴って複合体どうしが会合し、大きな分子量をもつ会合体を形成することが示された。この結果は、放射線によって切断されたゲノムDNAの2つの末端どうしをPprAが近づける役割をもつことを示唆し、PprAの機能発現において重要な知見であると考えられる。
安達 基泰; 玉田 太郎; 佐藤 勝也*; 鳴海 一成; 黒木 良太
no journal, ,
放射線抵抗性細菌デイノコッカスは、ヒト細胞の1000倍もの放射線抵抗性を示す。PprAはデイノコッカスより単離された新規なDNA修復促進蛋白質であり、高度放射線抵抗性において最も重要な役割を担っていることがこれまでに報告されている(1)。既にPprAは、別の新規な蛋白質(PprI)の発現誘導下にあること,2本鎖DNAをつなぐ作用(リガーゼ活性)を促進すること、及びDNAに結合能を有することが判明している。本研究では、PprAの構造と機能の関係を解明し、PprAの医療や産業への応用を推進するために、原子分解能レベルの詳細な立体構造解析を目的として研究を推進している。まず、大腸菌発現系を用いて得られたPprAの精製条件の検討を行い、大量調製する方法を確立した。この試料を用いて、結晶化条件をスクリーニングしたところ、微少なPprA単体の結晶化に成功した。PprAとDNAの結合をさまざまな角度から検討した結果、PprAとDNAの複合体において、PprAは1分子のDNA(pUC19,2686bp)に少なくとも280分子結合できることを明らかにした。また、2本鎖DNAの末端あるいはその単鎖部分に切断がある(ニックがある)場合、複数のDNA分子が結合して複合体を形成する可能性を示した。これらのことは、PprAの会合構造と機能発現に関係する重要な知見である。
奥津 賢一*; 木野 康志*; 中島 良太*; 宮下 湖南*; 安田 和弘*; 山下 琢磨*; 岡田 信二*; 佐藤 元泰*; 岡 壽崇; 河村 成肇*; et al.
no journal, ,
ミュオン触媒核融合(CF)は、負ミュオンがミュオン分子ddを形成し分子内で核融合反応(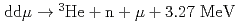 もしくは
もしくは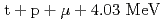 )を起こし、これを次々と繰り返す反応である。我々はCFの素過程の研究のため、固体水素標的を真空槽中に作製し基礎実験を行っている。本発表では、固体水素標的の形状、状態などの観測の他にCF実験での実用性などについて報告する。
)を起こし、これを次々と繰り返す反応である。我々はCFの素過程の研究のため、固体水素標的を真空槽中に作製し基礎実験を行っている。本発表では、固体水素標的の形状、状態などの観測の他にCF実験での実用性などについて報告する。
小西 蓮*; 奥津 賢一*; 木野 康志*; 佐々木 喬祐*; 中島 良太*; 山下 琢磨*; 宮下 湖南*; 安田 和弘*; 岡田 信二*; 佐藤 元泰*; et al.
no journal, ,
負ミュオンが重水素分子イオン(D)様のミュオン分子ddを作ると、核間距離が小さくなり、核間の波動関数がクーロン障壁をトンネル効果ですり抜けて、強い相互作用が働く距離で有意な値を持ち、重なり、その結果としてミュオン分子内で核融合が起こる。核融合後に放出されるミュオンは、高品質なミュオンビーム源として期待されているが、その挙動が不明であった。そこで、本研究ではPHITSコードを用いて、超低速ミュオンの固体水素薄膜中における振る舞いを調べた。厚さ0.1 1mで変化させた固体水素に110keVの単色ペンシルミュオンビームを打ち込んだところ、10keVの場合、1mではほとんどのミュオンが停止し、厚さ0.5mで70%、厚さ0.4mより薄いところでは
1mで変化させた固体水素に110keVの単色ペンシルミュオンビームを打ち込んだところ、10keVの場合、1mではほとんどのミュオンが停止し、厚さ0.5mで70%、厚さ0.4mより薄いところでは 99%のミュオンが固体水素薄膜を通過するなどがわかった。
99%のミュオンが固体水素薄膜を通過するなどがわかった。
