


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
岡村 正愛*; 中山 真義*; 梅基 直行*; Cano, E. A.*; 長谷 純宏; 西崎 雄三*; 佐々木 伸大*; 小関 良宏*
Euphytica, 191(1), p.45 - 56, 2013/05
被引用回数:28 パーセンタイル:80.53(Agronomy)液胞内アントシアニン凝集(AVI)によって特殊なdusky花色を発色するカーネーション系統がわずかではあるが知られている。特殊花色は非アシル化アントシアニンを含むAVIの存在と強く相関のある単一劣勢因子によって支配されることが遺伝様式から示唆されている。われわれは、カーネーションの特殊花色を多様化するため、交配育種によって新奇なメタリック花色を発色する青色系カーネーションを作出した。さらに、この系統にイオンビームを照射することにより、赤,赤紫及びクリムゾン系のメタリック花色系統を作出した。すべてのメタリック花色系統はアントシアニンマリル基転移酵素の転写産物を持たなかった。また、dusky花色のカーネーションと比べて、強く凝集したAVIと透明の液胞を持つ向軸側の表皮細胞を有していた。これらの結果から、(1)AVIを生成する因子はアントシアニンマリル基転移酵素の不活化によること、(2)向軸側表皮細胞のAVIがメタリック調花色を発色すること、(3)イオンビームがアントシアニン構造とAVIの凝集程度を変化することにより、メタリック花色の拡大に有用であることを示した。
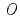 -methyltransferase involved in flower coloration
-methyltransferase involved in flower coloration秋田 祐介; 北村 智; 長谷 純宏; 鳴海 一成; 石坂 宏*; 近藤 恵美子*; 亀有 直子*; 中山 真義*; 谷川 奈津*; 森田 裕将*; et al.
Planta, 234(6), p.1127 - 1136, 2011/12
被引用回数:41 パーセンタイル:75.95(Plant Sciences)Anthocyanin -methyltransferase (OMT) is one of the key enzymes for anthocyanin modification and flower pigmentation. We previously bred a novel red-purple-flowered fragrant cyclamen (KMrp) from the purple-flowered fragrant cyclamen "Kaori-no-mai" (KM) by ion-beam irradiation. Since the major anthocyanins in KMrp and KM petals were delphinidin 3,5-diglucoside and malvidin 3,5-diglucoside, respectively, inactivation of a methylation step in the anthocyanin biosynthetic pathway was indicated in KMrp. We isolated and compared 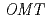 genes expressed in KM and KMrp petals. RT-PCR analysis revealed that
genes expressed in KM and KMrp petals. RT-PCR analysis revealed that 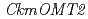 was expressed in the petals of KM but not in KMrp. Three additional
was expressed in the petals of KM but not in KMrp. Three additional 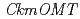 s with identical sequences were expressed in petals of both KM and KMrp. Genomic PCR analysis revealed that was not amplified from the KMrp genome, indicating that ion-beam irradiation caused a loss of the entire region in KMrp. In vitro enzyme assay demonstrated that CkmOMT2 catalyzes the 3' or 3',5' -methylation of the B-ring of anthocyanin substrates. These results suggest that CkmOMT2 is functional for anthocyanin methylation, and defective expression of is responsible for changes in anthocyanin composition and flower coloration in KMrp.
s with identical sequences were expressed in petals of both KM and KMrp. Genomic PCR analysis revealed that was not amplified from the KMrp genome, indicating that ion-beam irradiation caused a loss of the entire region in KMrp. In vitro enzyme assay demonstrated that CkmOMT2 catalyzes the 3' or 3',5' -methylation of the B-ring of anthocyanin substrates. These results suggest that CkmOMT2 is functional for anthocyanin methylation, and defective expression of is responsible for changes in anthocyanin composition and flower coloration in KMrp.
近藤 恵美子*; 中山 真義*; 亀有 直子*; 谷川 奈津*; 森田 裕将*; 秋田 祐介; 長谷 純宏; 田中 淳; 石坂 宏*
JAEA-Review 2010-065, JAEA Takasaki Annual Report 2009, P. 65, 2011/01
埼玉県では、シクラメンの栽培種と芳香性野生種の種間交雑と染色体倍加により、バラやスズランと同様の香気成分を持つ複二倍体の芳香性シクラメンを育成した。しかし、芳香性シクラメンは色や形が限定されているため、幅広い消費者の要望に応じるために、イオンビームによる突然変異育種を試みた。芳香性シクラメン「香りの舞い」に炭素イオンビームを2Gy照射して得られたM2個体の中から、花色変異体が得られた。「香りの舞い」の主要色素がマルビジン3.5-ジグルコサイドであるのに対して、変異体の主要色素はシクラメンでは今までに見いだされていないデルフィニジン3,5-ジグルコサイドであった。以上の結果は、変異体では、デルフィニジンの3'位及び5'位の水酸基のメチル化酵素をコードする遺伝子が、イオンビームの影響を受け、マルビジンへの代謝が阻害されていることを示唆している。
秋田 祐介; 石坂 宏*; 中山 真義*; 島田 明彦; 北村 智; 長谷 純宏; 鳴海 一成; 田中 淳
Journal of Horticultural Science & Biotechnology, 85(5), p.437 - 443, 2010/09
シクラメンの花色合成遺伝子に関する解析のために、紫色の花を有するシクラメン野生種(,gra6)とその白花変異体(gra50)を用いて、花色成分とその合成にかかわる遺伝子群の比較解析を行った。gra6の花の色素は、アントシアニンのマルビジン3,5ジグルコシドであった。一方、gra50の花ではアントシアニンが確認されず、その前駆物質であるフラボノールの蓄積が確認された。花色合成にかかわる酵素遺伝子群をgra6より単離し、その発現をgra6とgra50で解析したところ、二つのdihydroflavonol-4-reductase(DFR)遺伝子(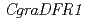 ,
, 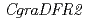 )のうち、の発現がgra50で減少していた。を含む他の遺伝子群には大差がみられなかったことから、がにおける花色に重要であり、その発現抑制によってアントシアニンの合成が進まず、白花に変異したと考えられた。の発現が減少した理由として、遺伝子発現にかかわるプロモーター領域の変異が大きな候補として挙げられた。本研究は、シクラメン花色合成に関する選抜マーカーとして
)のうち、の発現がgra50で減少していた。を含む他の遺伝子群には大差がみられなかったことから、がにおける花色に重要であり、その発現抑制によってアントシアニンの合成が進まず、白花に変異したと考えられた。の発現が減少した理由として、遺伝子発現にかかわるプロモーター領域の変異が大きな候補として挙げられた。本研究は、シクラメン花色合成に関する選抜マーカーとして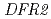 遺伝子が一つの候補であることを示した。
遺伝子が一つの候補であることを示した。
 spp., generated by ion-beam irradiation
spp., generated by ion-beam irradiation近藤 恵美子*; 中山 真義*; 亀有 直子*; 谷川 奈津*; 森田 裕将*; 秋田 祐介; 長谷 純宏; 田中 淳; 石坂 宏*
Plant Biotechnology, 26(5), p.565 - 569, 2009/01
被引用回数:33 パーセンタイル:66.84(Biotechnology & Applied Microbiology)香りシクラメンシリーズのバリエーションの増加のために、紫の花が咲き、主要アントシアニンとしてマルビジン3,5-ジグルコサイドを含む香りシクラメン品種(
 )「香りの舞」の黄化葉柄に320MeVの炭素イオンビームを0
)「香りの舞」の黄化葉柄に320MeVの炭素イオンビームを0 16Gyの強さで照射した。M1植物の自家受粉により得られたM2植物から変異体が選抜された。そのうち、2Gyを照射して得られたM2植物68個体中9個体は、これまでの紫の花と葉は異なる赤紫の花が咲いた。その花の色素を抽出し液体クロマトグラフィー(HPLC)で解析したところ、その主要アントシアニンは、デルフィニジン3,5-ジグルコサイドであった。花の形態や香り成分等、花の色以外の要素はこれまでのシクラメン品種とは大差がなかったことから、この変異体は花の色素合成にかかわる遺伝子にのみ変異が生じたと考えられる。デルフィニジン3,5-ジグルコサイドを主な色素として持つシクラメンはこれまで報告されておらず、この変異個体は商業的な価値があるだけでなく、シクラメンの貴重な遺伝子資源としても有用である。
16Gyの強さで照射した。M1植物の自家受粉により得られたM2植物から変異体が選抜された。そのうち、2Gyを照射して得られたM2植物68個体中9個体は、これまでの紫の花と葉は異なる赤紫の花が咲いた。その花の色素を抽出し液体クロマトグラフィー(HPLC)で解析したところ、その主要アントシアニンは、デルフィニジン3,5-ジグルコサイドであった。花の形態や香り成分等、花の色以外の要素はこれまでのシクラメン品種とは大差がなかったことから、この変異体は花の色素合成にかかわる遺伝子にのみ変異が生じたと考えられる。デルフィニジン3,5-ジグルコサイドを主な色素として持つシクラメンはこれまで報告されておらず、この変異個体は商業的な価値があるだけでなく、シクラメンの貴重な遺伝子資源としても有用である。
秋田 祐介; 石坂 宏*; 島田 明彦; 中山 真義*; 北村 智; 長谷 純宏; 田中 淳; 鳴海 一成
no journal, ,
シクラメンの花色とその生合成に関する遺伝子群との関連を調べるために、紫色の花色を有するシクラメン野生種(C. graecum, 2n=4x=84)とその花色が白色である変異体を用いて、それぞれの花弁における色素分析とアントシアニン生合成に関与する遺伝子群の比較解析を行った。HPLCの結果、野生種(gra6)のアントシアニンはマルビジン-3,5-ジグルコシド(Mv3,5dG)であった。一方、白色変異体(gra50)ではアントシアニンがほとんど確認されず、フラボノールが、gra6と比較すると増加していた。このgra50における花弁の色素減少の原因因子を探るため、gra6よりアントシアニン合成関連遺伝子群の単離を試みた。その結果、フラボノール合成以降に関与する酵素遺伝子のホモログとして、フラボノイド3'5'-水酸化酵素を1種類(CgF3'5'H),ジヒドロフラボノール4-還元酵素を2種類(CgDFR1, CgDFR2),アントシアニジン合成酵素を1種類(CgANS)それぞれ単離した。それぞれの遺伝子の発現を解析したところ、CgF3'5'H, CgDFR1, CgANSの発現は、gra6, gra50のスリップでともに強い発現がみられ、2種間で大差がなかった。一方、CgDFR2の発現はgra6で強く確認されたが、gra50ではその発現は極端に減少していた。
近藤 恵美子*; 亀有 直子*; 中山 真義*; 秋田 祐介; 長谷 純宏; 谷川 奈津*; 森田 裕将*; 石坂 宏*
no journal, ,
芳香シクラメン「麗しの香り」の半数体にイオンビーム照射を行い、選抜したサーモンピンクの変異体(ion3)にイオンビーム再照射を行い、変異体の作出を試みた。照射集団から形態観察により個体を選抜し、それらの中から、さらに花弁の縁に欠刻が入った変異体(ion243),花色のサーモンピンクが濃くなった変異体(ion245),アイの部分が欠失し、花弁全体がサーモンピンクになった変異体(ion246, ion255)を選抜した。ion3と変異体の花弁の大きさを比較すると、変異体は縦長,横長ともion3に比べて小さかった。スリップの主要アントシアニンは、シアニジン3,5-ジグルコサイド,ペオニジン3,5-ジグルコサイド,マルビジン3,5-ジグルコサイド(Mv3,5dG)であった。ion3及び変異体のアイの色は濃い赤紫色で、主要アントシアニンはMv3,5dGであった。アントシアンニン濃度をion3と変異体とで比較すると、スリップではion3に比べてion243, ion245, ion246で高く、ion255では低かった。アイのアントシアニン濃度はion3に比べてion243, ion245で高く、スリップのアントシアニン濃度と同様の傾向が認められた。特に、花色が濃いと観察されたion245では、スリップ及びアイのアントシアニン濃度がion3の約2倍であることが明らかとなった。
and its white-flowered mutant秋田 祐介; 石坂 宏*; 中山 真義*; 島田 明彦; 北村 智; 長谷 純宏; 田中 淳; 鳴海 一成
no journal, ,
To reveal the relationship between floral pigmentation and flavonoid biosynthesis genes in cyclamen ( species), we analyzed flower pigments from wild-type cyclamen ( ) and its white-flowered mutant. Analysis with high performance liquid chromatography (HPLC) showed that major anthocyanins of the wild-type petal were malvidin 3,5-diglucosides. The white-flowered mutant possessed smaller amount of anthocyanins but higher amount of flavonols than the wild-type, suggesting the change of metabolic flow by disruption of anthocyanin biosynthesis. By degenerate-PCR with total RNAs from wild-type petals, we isolated some flavonoid biosynthesis gene. RT-PCR using a specific primer set for each gene demonstrated that the expression of decreased in white-flowered mutant compared with wild-type, whereas the expressions of other genes did not appear to differ greatly. The genomic construction of was not different between wild-type and white-flowered mutant, inferring that reduced expression of correlates with white-flowered mutation. These results suggest that is involved in anthocyanin biosynthesis pathway in , and the white-flowered mutation could be caused by transcriptional regulation of gene.
) and its white-flowered mutant. Analysis with high performance liquid chromatography (HPLC) showed that major anthocyanins of the wild-type petal were malvidin 3,5-diglucosides. The white-flowered mutant possessed smaller amount of anthocyanins but higher amount of flavonols than the wild-type, suggesting the change of metabolic flow by disruption of anthocyanin biosynthesis. By degenerate-PCR with total RNAs from wild-type petals, we isolated some flavonoid biosynthesis gene. RT-PCR using a specific primer set for each gene demonstrated that the expression of decreased in white-flowered mutant compared with wild-type, whereas the expressions of other genes did not appear to differ greatly. The genomic construction of was not different between wild-type and white-flowered mutant, inferring that reduced expression of correlates with white-flowered mutation. These results suggest that is involved in anthocyanin biosynthesis pathway in , and the white-flowered mutation could be caused by transcriptional regulation of gene.
秋田 祐介; 石坂 宏*; 中山 真義*; 北村 智; 長谷 純宏; 田中 淳; 鳴海 一成
no journal, ,
イオンビーム照射による植物の変異体作製は、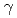 線照射に比べて突然変異率が高く、新品種の作出に直結しやすいことから、増加傾向にある。我が国で最も栽培されている鉢植え植物であるシクラメンは、花色のバリエーションが少なく、その広がりが求められている。そこでシクラメンのイオンビーム照射による新しい花色を持つ変異体作製が進められているが、シクラメンは種子から開花までの期間が長く、有望な変異個体の効率的な選抜には花色の選抜マーカーの開発が必要である。しかしながら、シクラメンの花色に関する知見は少なく、選抜マーカー開発のためには、花色とそれにかかわる遺伝子との相関関係を明らかにすることが必須である。そこでシクラメンの花色とその生合成に関する遺伝子群との関連を調べるために、紫色の花色を有するシクラメン野生種とその花色が白色である変異体を用いて、それぞれの花弁における色素分析と色素生合成に関与する遺伝子群の比較解析を行った。
線照射に比べて突然変異率が高く、新品種の作出に直結しやすいことから、増加傾向にある。我が国で最も栽培されている鉢植え植物であるシクラメンは、花色のバリエーションが少なく、その広がりが求められている。そこでシクラメンのイオンビーム照射による新しい花色を持つ変異体作製が進められているが、シクラメンは種子から開花までの期間が長く、有望な変異個体の効率的な選抜には花色の選抜マーカーの開発が必要である。しかしながら、シクラメンの花色に関する知見は少なく、選抜マーカー開発のためには、花色とそれにかかわる遺伝子との相関関係を明らかにすることが必須である。そこでシクラメンの花色とその生合成に関する遺伝子群との関連を調べるために、紫色の花色を有するシクラメン野生種とその花色が白色である変異体を用いて、それぞれの花弁における色素分析と色素生合成に関与する遺伝子群の比較解析を行った。
近藤 恵美子*; 中山 真義*; 亀有 直子*; 谷川 奈津*; 森田 裕将*; 秋田 祐介; 長谷 純宏; 田中 淳; 石坂 宏*
no journal, ,
芳香シクラメン(2n=4x=82)は花の色が紫とピンクに限られている。花色のバラエティーを広げるために、イオンビーム照射による突然変異育種を試みた。花色が紫色の芳香シクラメン品種"香りの舞い"を暗黒下で培養し、もやし状の黄化葉柄を育成した。これに320MeVの炭素イオンビームを照射し再分化個体を得た。この後代の種子を育成し形質調査を行ったところ、花の色が赤紫色に変異した個体が得られた。この変異体の色素を分析したところ、"香りの舞い"の主要色素であるマルビジン3,5ジグルコシドとは異なる、デルフィニジン3,5ジグルコシドという色素が主要色素として含まれていた。今回得られた突然変異体は赤紫色の新品種になり得るとともに、シクラメン属植物においてデルフィニジン系色素を持つ園芸品種及び野生種はないことから、育種における貴重な遺伝資源として利用できる。
近藤 恵美子*; 亀有 直子*; 中山 真義*; 栗原 康*; 秋田 祐介; 谷川 奈津*; 森田 裕将*; 長谷 純宏; 田中 淳; 石坂 宏*
no journal, ,
芳香シクラメンは花の色が紫とピンクに限られており、花色や花型の変化が乏しい。そこで、芳香シクラメン品種"麗しの香り"の半数体にイオンビーム照射を行い、サーモンピンクの変異体(ion3)を選抜した。一方、イオンビーム育種では照射個体に再度照射を行うことにより、さらに広い変異が得られる可能性がある。そこで、さらなる変異拡大を目指してion3にイオンビーム再照射を行い、変異体の作出を試みた。その結果、花弁の縁に欠刻が入った変異体や、ion3に比べて花色のサーモンピンクが濃くなった変異体、花の中心部にあるアイの部分が欠失し、花弁全体がサーモンピンクになった変異体を選抜した。このことは、イオンビームの再照射によって変異頻度が高まっていることを示している。
岡村 正愛*; 長谷 純宏; 梅基 直行*; 中山 真義*; 谷川 奈津*; 鳴海 一成; 田中 淳
no journal, ,
特定の形質を狙ってその形質が変化した個体を多く得ることができれば、画期的な技術となる。春先に白いはずの花に赤い色素がさすことがよくある。これは光合成が増す一方で寒さも残り代謝産物が色素合成にまわるためと推定した。そこで糖処理により色素が誘導され状態でイオンビームを照射すれば、色素遺伝子が発現した状態でイオンビームが細胞に当てることができ色素遺伝子の変異が多く得られると洞察した。ペチュニア幼苗にショ糖溶液を与え茎や芽に色素を誘導した後に炭素イオンビームを照射した。この種子を播種し、葉緑素変異体及び花色変異体が分離したM2系統の頻度を調査した。独立した3回の試験のいずれでも、葉緑素変異の頻度は蔗糖処理に有無によらず同じであったのに対して、花色の変異は蔗糖処理区の頻度が無処理句に比較して2倍以上高く、照射前の蔗糖処理によって花色変異体の獲得頻度が有意に向上することが示された。以上の試験により、突然変異の方向性をある程度制御できることが示唆された。
 )の半数体へのイオンビーム照射によって得られた赤紫色変異体の色素と香気成分の解析
)の半数体へのイオンビーム照射によって得られた赤紫色変異体の色素と香気成分の解析亀有 直子*; 中山 真義*; 近藤 恵美子*; 栗原 康*; 秋田 祐介; 長谷 純宏; 谷川 奈津*; 森田 裕将*; 田中 淳; 石坂 宏*
no journal, ,
芳香シクラメン"香りの舞い"( )の花色を変えるためにイオンビーム照射による突然変異育種を試みた。変異体を効率よく選抜するためには半数体の利用が有効であるため、葯培養により半数体を育成した。その上で半数体へのイオンビーム照射により変異体を獲得し、花色及び香気成分の分析を行った。77個の葯を培養した結果、123個の幼植物体が再分化した。この中から54個の幼植物体を用いてDAPI蛍光強度を測定した結果、53個体が指標個体と同じゲノムサイズを持つ半数体であると判断された。確認した個体を照射材料として、2,863の切片にイオンビーム照射を行い、照射集団の中から形態観察により花弁の色が赤紫色となった変異体を選抜した。赤紫色変異体は、赤紫色の花と半数体と同様の紫色の花が同時に咲くキメラであった。花弁の主要色素はスリップ,アイで共通であり、半数体がマルビジン3,5-ジグルコサイド、変異体がマルビジン3-グルコサイドであった。半数体と変異体の花弁の形態及び香気成分に大きな変化は見られなかった。以上のような特徴を持った赤紫色変異体は、アントシアニンの結合糖が欠損したことにより得られたと考えられた。
)へのイオンビーム照射によって得られた赤紫色変異体の色素と香気成分の解析近藤 恵美子*; 中山 真義*; 亀有 直子*; 栗原 康*; 谷川 奈津*; 森田 裕将*; 秋田 祐介; 長谷 純宏; 田中 淳; 石坂 宏*
no journal, ,
種間交雑により作出した芳香シクラメン( )の変異を拡大するためにイオンビーム照射を行いM2集団より変異体を選抜した。この変異体の色素と香気成分を解析する。222個のM1個体に形態的変異は認められず、そこから271個のM2開花植物体を育成した。その中で、2Gy照射の68個体から9個の花色突然変異体を選抜した。これらの突然変異体は、花の形、大きさ、葉色及び香りは変化していなかったが、花の色が赤紫色に変異していた。"香りの舞い"のスリップの主要色素はマルビジン3,5-ジグルコシドであり、花色突然変異体9個体の主要色素はデルフィニジン3,5-ジグルコシドであった。
)へのイオンビーム照射によって得られた白色変異体の色素と香気成分の解析近藤 恵美子*; 中山 真義*; 亀有 直子*; 栗原 康*; 谷川 奈津*; 森田 裕将*; 秋田 祐介; 長谷 純宏; 田中 淳; 石坂 宏*
no journal, ,
種間交雑により作出した芳香シクラメン( )"孤高の香り"の変異を拡大するためにイオンビーム照射による変異体の作出を行い、得られた変異体の解析を行う。239の葉片にイオンビームを照射し、100個の植物体を再生させた。そのうち、34個のM1個体が開花したが、形態的変異は認められず、すべての形質が"孤高の香り"と同様であった。M1個体の自家受粉により74粒のM2種子を採取し、71個の植物体を育成した。その中から6個体の白色の花色突然変異体を選抜した。"孤高の香り"のアイとスリップの主要アントシアニンはマルビジン3,5-ジグルコサイド(Mv3,5-dG)であり、フラボノールとしてケルセチン(Que)とケンフェロール(Kae)が検出された。白色の変異体からMv3,5-dGは検出されず、QueとKaeのみが検出された。変異体によってQueとKaeの量に違いがあり、"孤高の香り"よりも有意に増加した変異体が見られた。
近藤 恵美子*; 亀有 直子*; 石坂 宏*; 中山 真義*; 谷川 奈津*; 森田 裕将*; 秋田 祐介; 長谷 純宏; 田中 淳
no journal, ,
埼玉県では、シクラメンの栽培種と芳香性野生種の種間交雑と染色体倍加により、バラやスズランと同様の香気成分を持つ複二倍体の芳香性シクラメンを育成した。しかし、芳香性シクラメンは色や形が限定されているため、幅広い消費者の要望に応じるために、イオンビームによる突然変異育種を試みた。芳香性シクラメン「香りの舞い」に炭素イオンビームを2Gy照射して得られたM2個体の中から、花色変異体が得られた。「香りの舞い」の主要色素がマルビジン3.5-ジグルコサイドであるのに対して、変異体の主要色素はシクラメンでは今までに見いだされていないデルフィニジン3,5-ジグルコサイドであった。以上の結果は、変異体では、デルフィニジンの3'位及び5'位の水酸基のメチル化酵素をコードする遺伝子が、イオンビームの影響を受け、マルビジンへの代謝が阻害されていることを示唆している。
)へのイオンビーム照射によって得られた白色変異体の香気成分の分析亀有 直子*; 大久保 直美*; 近藤 恵美子*; 中山 真義*; 秋田 祐介; 長谷 純宏; 谷川 奈津*; 森田 裕将*; 田中 淳; 石坂 宏*
no journal, ,
芳香シクラメン"孤高の香り"の変異を拡大するためにイオンビーム照射を行ったところ、M2集団から白色の変異体が得られた。これまでに、変異体では、花弁においてフラボノールのみが検出されアントシアニンは含まれていないこと、"孤高の香り"の主要香気成分であった芳香族化合物が検出されなかったことを報告した。アントシアニンと芳香族化合物は前駆体が共通であることから、花弁色素と香気成分の両方に変化が起きたことが推察されたため、本研究では変異体の香気成分に関する詳細な情報を得ることを目的として、"孤高の香り"及び白色変異体の花の発散成分及び内生成分の定量分析を行った。
)の半数体へのイオンビーム再照射による倍加半数体の作出近藤 恵美子*; 亀有 直子*; 北村 智; 秋田 祐介; 長谷 純宏; 田中 淳; 中山 真義*; 谷川 奈津*; 森田 裕将*; 石坂 宏*
no journal, ,
これまでに、芳香シクラメン()"麗しの香り"(複二倍体,花色:ピンク)の半数体にイオンビーム照射を行い、サーモンピンクの変異体(ion3)を選抜し、さらに、ion3へのイオンビーム再照射により新たな変異体(不稔)を得た。本研究ではion3の黄化葉柄へのイオンビーム再照射と培養により稔性を回復した個体(倍加半数体)を獲得したので報告する。この倍加半数体は大きさがion3よりも大きかったが、花色の成分には大差がなく、種子培養によって個体を増殖させることが可能となり、新品種作出に大きな期待が持てる。
)へのイオンビーム照射によって得られた変異体の色素の解析近藤 恵美子*; 中山 真義*; 亀有 直子*; 谷川 奈津*; 森田 裕将*; 北村 智; 秋田 祐介; 長谷 純宏; 田中 淳; 石坂 宏*
no journal, ,
種間交雑により作出した芳香シクラメン"香りの舞い"()の変異を拡大するためにイオンビーム照射を行った。その結果、"香りの舞い"の花色は紫色であるが、320MeV炭素イオンビーム1Gy及び2Gy照射区のM2集団から、それぞれ8個及び11個の赤紫色の花色変異体が得られた。1Gy及び2Gy照射由来の変異体のスリップは赤紫色、アイは濃赤紫色であり、両変異体の花色による識別は困難であった。"香りの舞い"のスリップ及びアイの主要アントシアニンはマルビジン3,5-ジグルコシド(Mv3,5-dG)であった。1Gy照射由来の8個体の変異体のスリップとアイから大量のマルビジン3-グルコシド(Mv3-G)が検出されたが、そのほかに微量のデルフィニジン3-グルコシド(Dp3-G)とペチュニジン3-グルコシド(Pt3-G)が検出された。2Gy照射由来の11個の変異体のスリップ及びアイからデルフィニジン3,5-ジグルコシド(Dp3,5-dG)が検出された。"香りの舞い"と変異体のアントシアニン濃度はスリップよりもアイで高くなったことから、スリップとアイの色調の違いはアントシアニン濃度によると考えられる。これらの変異体は新品種候補としてだけでなく、育種素材としても期待される。
岡村 正愛*; 中山 真義*; 長谷 純宏
no journal, ,
ほとんどのカーネーションの花弁細胞はリンゴ酸でアシル化されたアントシアニンを含有するが、メタリック調の輝きを持つ一部の特殊花色系統では、アシル化されていないアントシアニンを含有することが知られている。通常、細胞の液胞内全体に分布するアントシアニンが、特殊花色系統では、液胞内で凝集体を形成することによってメタリック調の花色を示すと考えられている。われわれは、アントシアニンをアシル化する酵素遺伝子を指標として、多数の通常花色系統からアシル化能をヘテロで欠損した系統を複数見いだし、それらを交配することによって青紫色のメタリック色調の特殊花色系統の作出に成功した。さらに青紫メタリック系統へのイオンビーム照射によって色調のバリエーション拡大を試み、赤紫メタリック系統,銅赤メタリック系統並びに赤メタリック系統を獲得することに成功した。これらの変異系統が含有するアントシアニンを分析した結果、色素量,糖修飾及び凝集性に変化が見られ、これらが色調の違いを生み出した原因であると考えられた。
