


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
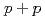 collisions at
collisions at 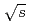 = 200 GeV
= 200 GeVAdare, A.*; Afanasiev, S.*; Aidala, C.*; Ajitanand, N. N.*; Akiba, Y.*; Al-Bataineh, H.*; Alexander, J.*; Aoki, K.*; Aphecetche, L.*; Armendariz, R.*; et al.
Physical Review D, 84(1), p.012006_1 - 012006_18, 2011/07
被引用回数:29 パーセンタイル:72.31(Astronomy & Astrophysics)重心エネルギー200GeVでの縦偏極陽子陽子衝突からのジェット生成のイベント構造と二重非対称(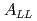 )について報告する。光子と荷電粒子がPHENIX実験で測定され、イベント構造がPHYTIAイベント生成コードの結果と比較された。再構成されたジェットの生成率は2次までの摂動QCDの計算で十分再現される。測定されたは、一番低い横運動量で-0.0014
)について報告する。光子と荷電粒子がPHENIX実験で測定され、イベント構造がPHYTIAイベント生成コードの結果と比較された。再構成されたジェットの生成率は2次までの摂動QCDの計算で十分再現される。測定されたは、一番低い横運動量で-0.0014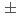 0.0037、一番高い横運動量で-0.01810.0282であった。このの結果を幾つかの
0.0037、一番高い横運動量で-0.01810.0282であった。このの結果を幾つかの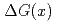 の分布を仮定した理論予想と比較する。
の分布を仮定した理論予想と比較する。
collisions at = 200 and 62.4 GeVAdare, A.*; Afanasiev, S.*; Aidala, C.*; Ajitanand, N. N.*; 秋葉 康之*; Al-Bataineh, H.*; Alexander, J.*; 青木 和也*; Aphecetche, L.*; Armendariz, R.*; et al.
Physical Review C, 83(6), p.064903_1 - 064903_29, 2011/06
被引用回数:184 パーセンタイル:99.44(Physics, Nuclear)200GeVと62.4GeVでの陽子陽子の中心衝突からの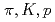 の横運動量分布及び収量をRHICのPHENIX実験によって測定した。それぞれエネルギーでの逆スロープパラメーター、平均横運動量及び単位rapidityあたりの収量を求め、異なるエネルギーでの他の測定結果と比較する。また
の横運動量分布及び収量をRHICのPHENIX実験によって測定した。それぞれエネルギーでの逆スロープパラメーター、平均横運動量及び単位rapidityあたりの収量を求め、異なるエネルギーでの他の測定結果と比較する。また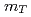 や
や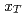 スケーリングのようなスケーリングについて示して陽子陽子衝突における粒子生成メカニズムについて議論する。さらに測定したスペクトルを二次の摂動QCDの計算と比較する。
スケーリングのようなスケーリングについて示して陽子陽子衝突における粒子生成メカニズムについて議論する。さらに測定したスペクトルを二次の摂動QCDの計算と比較する。
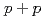 and Au+Au collisions at
and Au+Au collisions at 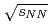 = 200 GeV
= 200 GeVAdare, A.*; Afanasiev, S.*; Aidala, C.*; Ajitanand, N. N.*; 秋葉 康之*; Al-Bataineh, H.*; Alexander, J.*; 青木 和也*; Aphecetche, L.*; Aramaki, Y.*; et al.
Physical Review C, 83(4), p.044912_1 - 044912_16, 2011/04
被引用回数:8 パーセンタイル:49.7(Physics, Nuclear)重いフレーバーのメソンの崩壊からの電子の測定は、このメソンの収量が金金衝突では陽子陽子に比べて抑制されていることを示している。われわれはこの研究をさらに進めて二つの粒子の相関、つまり重いフレーバーメソンの崩壊からの電子と、もう一つの重いフレーバーメソンあるいはジェットの破片からの荷電ハドロン、の相関を調べた。この測定は重いクォークとクォークグルオン物質の相互作用についてのより詳しい情報を与えるものである。われわれは特に金金衝突では陽子陽子に比べて反対側のジェットの形と収量が変化していることを見いだした。
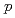 + collisions at = 200 GeV and scaling properties of hadron production
+ collisions at = 200 GeV and scaling properties of hadron productionAdare, A.*; Afanasiev, S.*; Aidala, C.*; Ajitanand, N. N.*; Akiba, Y.*; Al-Bataineh, H.*; Alexander, J.*; Aoki, K.*; Aphecetche, L.*; Armendariz, R.*; et al.
Physical Review D, 83(5), p.052004_1 - 052004_26, 2011/03
被引用回数:175 パーセンタイル:98.48(Astronomy & Astrophysics)RHIC-PHENIX実験で重心エネルギー200GeVの陽子陽子衝突からの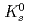 ,
, 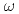 ,
, 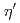 と
と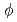 中間子生成の微分断面積を測定した。これらハドロンの横運動量分布のスペクトルの形はたった二つのパラメーター、
中間子生成の微分断面積を測定した。これらハドロンの横運動量分布のスペクトルの形はたった二つのパラメーター、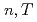 、のTsallis分布関数でよく記述できる。これらのパラメーターはそれぞれ高い横運動量と低い横運動量の領域のスペクトルを決めている。これらの分布をフィットして得られた積分された不変断面積はこれまで測定されたデータ及び統計モデルの予言と一致している。
、のTsallis分布関数でよく記述できる。これらのパラメーターはそれぞれ高い横運動量と低い横運動量の領域のスペクトルを決めている。これらの分布をフィットして得られた積分された不変断面積はこれまで測定されたデータ及び統計モデルの予言と一致している。
草野 俊胤; 小森 芳昭; 岩永 雅之; 都所 昭雄; 小松 久人; 舛井 仁一; 三浦 信之*; Lovett, J. E.*
PNC TN8410 86-52, 66 Pages, 1987/01
再処理工場の保障措置の効果を高めることを目的として構築されたニア・リアル・タイム計量管理(NRTA)の実証試験を動燃東海再処理工場で実施した。 実証試験は1980年のC-1キャンペーンから開始し,その後データの蓄積のため継続して実施してきている。特に,1985年の後半の85-2キャンペーンでは,より実際的な形で実証試験を行うために査察行為を模擬した形で取り入れ,IAEAへの支援プログラムの一つとして実施した。 本報告書は,C-1キャンペーンから85-2キャンペーンまでに実施した実証試験により得られたデータおよびNRTAの効果などに関する知見をとりまとめたものである。 なお,本報告書はIAEAと平行して,2機関が同時に発行するものである。
Russo, P. A.*; 朝倉 祥郎*; Hsue, S. T.*; 近藤 勲*; Sprinkle, J. K. Jr.*; 舛井 仁一*; Johnson, S. S.*; 庄司 和弘*
PNC TN841 82-10, 76 Pages, 1982/08
None
朝倉 祥郎*; 宮原 顕治; 後藤 憲一*; 舛井 仁一*; 近藤 勲*; 寺門 茂*; 幸 文教*; 鎌田 正行
PNC TN852 81-18, 266 Pages, 1981/05
再処理工場における各工程に関する計量分析方法についてマニュアル化したものである。本編は特に計量管理分析関係の分析方法について編集したもので,再処理工程試料中のウラン,プルトニウム,全アルファ,密度,不純物の分析操作手順について記述した。
宮原 顕治; 朝倉 祥郎*; 後藤 憲一*; 舛井 仁一*; 近藤 勲*; 寺門 茂*; 鎌田 正行; 佐藤 悦朗*
PNC TN852 81-15, 46 Pages, 1981/04
再処理工場分析課に於いて,計量管理分析をするために必要なプルトニウム(一次)標準試料,ウラン(一次)標準試料,U-Pu混合(一次)標準試料の調製を実施したので報告する。
中村 久*; 舛井 仁一*; 坪谷 隆夫*
PNC TN841 77-59, 34 Pages, 1977/11
核燃料に含まれる核物質の分析技術は従来有効な手法として破壊分析法があるが、核燃料サイクルの過程には破壊分析の適用が難しいことも多い。特に各施設で発生する核物質を含んだスクラップや廃棄物は破壊分析を適用することが難しく、核物質の計量管理や保障措置上より簡便で適用性の広い非破壊分析技術の開発が必要とされている。アクティブ中性子核分裂法は一般にパッシブ法より感度が高く、高速中性子を用いれば透過性が高いので大きな試料や高濃縮の試料の分析が可能とされているが、この手法に関する我が国の報告例はほとんど無い。そのためにインテルコム・ラドテック社から中性子照射非破壊測定装置(ISAS)を購入し、アクティブ分析法による実験検討を行った。対象核物質として密封した濃縮ウラン、プルトニウムを用い、くり返し測定による計数値の再現性や日内、日間変動、幾何学的条件の影響、濃縮度の影響、等について検討した。また、模擬焼 灰、鉛や水、ポリエチレン、ビニルバック等の含水素物質を含む均質、非均質系試料を用いて実験を行いマトリックスの影響を検討した。その結果、核物質(UO/SUB2、PuO/SUB2粉末)およびこれらの核物質が均一化された状態のスクラップについては定量可能であることを確認した。
坪谷 隆夫*; 鎌田 正行; 和田 幸男; 舛井 仁一*; 山田 一夫*; 和田 勉*; 加藤木 賢; 高橋 信二*
PNC TN851 76-01, 42 Pages, 1976/01
TIDー7029(2nd edition)'Selected Measurement Methods for Plutonium and Unanium in the Nuclear Fuel Cycle, Second Edition(C,J,Rodden 編集)(1972)'の抜粋訳である。本訳著にはウランの湿式分析法、ウランおよびプルトニウムの同位体測定法(質量分析法、放射化学分析法)、同位体測定に必要な前処理技術、および同位体希釈質量分析法によるウランおよびプルトニウムの定量法を含んでいる。分析法毎に当分析所の手法に照らして訳注を付した。
増井 友美; 小泉 智; 橋本 竹治; 岩瀬 裕希; 敷中 一洋*; Kwon, H.*; 角五 彰*; Gong, J.*
no journal, ,
アクチン分子は生体内に最も豊富に存在する球状タンパク質であり、生体内のアクチン結合タンパク質とともにコンプレックスを形成することで高次構造を形成し、細胞運動を担うことが知られている。このため、細胞運動を理解するためにアクチン分子が形成するコンプレックスの構造の研究は精力的に進められ、蛍光標識したアクチン分子を用いて蛍光顕微鏡観測を中心に研究が進められている。生体内のアクチン濃度は高いが、蛍光顕微鏡観測では高アクチン濃度では背景光が蛍光ノイズとなるため高濃度での振る舞いに関しては不明な点が多いのが現状である。本研究では、中性子超小角散乱法を利用することで、顕微鏡実験より高濃度での実験を実現し、アクチンコンプレックスの構造を追跡した。その結果、顕微鏡実験の濃度ではコンプレックスを形成しない条件下でも、アクチン濃度を高くすることによりコンプレックスを形成することを明らかにした。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
アクチン分子は生体内に最も豊富に存在する球状タンパク質であり、生体内のアクチン結合タンパク質と結合することで高次構造を形成し細胞運動を担う。このため、細胞運動のメカニズムを明らかにするために、アクチン分子が形成するコンプレックスの構造の研究は精力的に進められてきた。従来のアクチンの研究は蛍光顕微鏡手法により行われ、低濃度でのアクチンの構造形成が明らかになってきた。しかしながら、高濃度のアクチンの観測は、蛍光による背景光がノイズとなるためその振る舞いはほとんど調べられていない。一方、生体内のアクチン濃度は高く、高濃度領域のアクチンの構造形成を調べることは、細胞運動のメカニズムを明らかにするうえで大変重要となる。そこで、本研究では中性子超小角散乱法を利用し、蛍光標識なしかつ高濃度といったより生体内に近い実験条件下でのアクチンコンプレックスの構造を追跡した。その結果、顕微鏡実験の濃度ではコンプレックスを形成しない条件でも、高濃度のアクチンではコンプレックスを形成すること、また、低濃度のアクチンのコンプレックス形成とは異なる経路で構造を形成していくことを明らかにした。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
フィラメントアクチンは、球状タンパク質であるアクチン分子が直線上に会合した細長い棒状分子集合体である。アクチンは非筋肉細胞中に存在し、その運動をバンドル構造やネットワーク構造などの高次構造の形成と解体によって制御している。この制御機構については、生化学的な研究では、生体内のアクチン結合タンパク質の多様性に起因するとされていたため、塩濃度による構造制御についてはほとんど検討がなされてこなかった。本研究では、アクチン結合タンパク質の代わりにポリカチオンを用い、フィラメントアクチンとポリカチオンの複合体に及ぼす塩濃度の効果を中性子小角散乱法により検討した。中性子超小角散乱法では、生体内に近い高濃度かつ溶液条件での実験が可能であり、本研究では、塩濃度変化による複合体構造の変化をナノメートルスケールからサブマイクロスケールにわたる空間領域で観察を行った。その結果、生体内の塩濃度に近い領域で、塩濃度をわずかに変化させることによってバンドル構造を解体することを明らかにした。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
アメーバ運動は、生体内の細胞骨格タンパク質であるアクチンがアクチン結合タンパク質を介して形成するバンドル,二次元ネットワーク,三次元ゲル構造の構築と消滅の制御によって生み出されている。本研究では、アクチン結合タンパク質のモデルとしてポリカチオンを用い、フィラメントアクチンとポリカチオンが形成する複合体構造を中性子超小角散乱法を用いて観察した。その結果、ポリカチオン濃度の増大によるバンドル内のフィラメント密度の増大、及び、塩濃度の増加によるバンドル構造の消滅を明らかにした。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
アメーバ運動は、生体内の細胞骨格タンパク質であるアクチンがアクチン結合タンパク質を介して形成するバンドル,二次元ネットワーク,三次元ゲル構造の構築と消滅の制御によって生み出されている。本研究では、アクチン結合タンパク質のモデルとしてカチオン性鎖状高分子を用い、フィラメントアクチンとカチオン性鎖状高分子が形成する複合体構造を中性子超小角散乱法を用いて観察した。その結果、塩濃度の増大によってネットワーク様構造からバンドル構造へと構造変化すること、さらに塩濃度が増大するとバンドル構造が消滅することを明らかにした。本研究の結果は、細胞運動における塩濃度の局所分布の重要性を示唆する。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
アメーバ運動は、生体内の細胞骨格タンパク質であるアクチンがアクチン結合タンパク質を介して形成するバンドル,二次元ネットワーク,三次元ゲル構造の構築と消滅の制御によって生み出されている。この構造の制御機構については、これまでアクチン結合タンパク質の多様性によって理解されてきた。一方、この系を物質科学的な性質に着目すると、アクチンは負電荷,アクチン結合タンパク質は正電荷を有する。また、生体内では塩濃度の不均一性が存在する。このため、塩濃度変化が構造形成に及ぼす効果を調べることは大変重要となる。そこで、本研究では、アクチン結合タンパク質のモデルとしてポリカチオンを用い、フィラメントアクチンとポリカチオンが形成する複合体構造に及ぼす塩濃度の効果を中性子超小角散乱法を用いて観察した。その結果、低塩濃度ではネットワーク様構造を形成し、塩濃度の増大によってバンドル構造が発達し、さらに塩濃度が上昇するとバンドル構造が消滅することを明らかにした。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
アメーバ運動は、生体内の細胞骨格タンパク質であるアクチンがアクチン結合タンパク質を介して形成するバンドル,二次元ネットワーク,三次元ゲル構造の構築と消滅の制御によって生み出されている。この構造の制御機構については、これまでアクチン結合タンパク質の多様性によって理解されてきた。一方、この系を物質科学的な性質に着目すると、アクチンは負電荷、アクチン結合タンパク質は正電荷を有する。また、生体内では塩濃度の不均一性が存在する。このため、塩濃度変化が構造形成に及ぼす効果を調べることは大変重要となる。そこで、本研究では、アクチン結合タンパク質のモデルとしてポリカチオンを用い、フィラメントアクチンとポリカチオンが形成する複合体構造に及ぼす塩濃度の効果を中性子超小角散乱法を用いて観察した。その結果、低塩濃度ではネットワーク様構造を形成し、塩濃度の増大によってバンドル構造が発達し、さらに塩濃度が上昇するとバンドル構造が消滅することを明らかにした。この構造変化が単価イオンによる静電遮蔽によって説明できることを明らかにした。
増井 友美; 敷中 一洋*; Kwon, H.*; 小泉 智; 橋本 竹治; 岩瀬 裕希; 角五 彰*; Gong, J.*
no journal, ,
細胞運動は、細胞骨格タンパク質であるアクチンがアクチン調節タンパク質を介して形成するバンドル,二次元ネットワーク,三次元ゲル構造の構築と消滅の制御によって生み出されている。これまでに、アクチンの負電荷,アクチン調節タンパク質の正電荷に着目し、アクチンとポリカチオンからなるモデル細胞骨格を構築すると、塩濃度変化によって構造変化が生じることが明らかにされてきた。しかしながら、顕微鏡分解能以下のナノメートルスケールの構造情報を得ることは困難であった。そこで、本研究では、蛍光顕微鏡と中性子小角散乱法を併用することで、溶液条件下でナノスケールからマクロメートルスケールに渡る階層構造の変化を明らかにすることを目的とし、4桁に渡る空間スケールの構造情報を定量的に得ることに成功した。得られた構造パラメータから、塩による遮蔽距離がアクチン間距離も大きい場合には、残存するアクチン間の斥力からコイル構造を形成すること、塩濃度の増大によってアクチン間の斥力が塩によって遮蔽されるとバンドルが成長し、さらなる塩濃度の増大は、アクチンとポリカチオン間の引力作用をも遮蔽することによって単一アクチンへと転移することを定性的に説明することができた。
増井 友美; 敷中 一洋*; 小泉 智; 橋本 竹治; 角五 彰*; Gong, J.*; Kwon, H.*
no journal, ,
アクチンとポリカチオン混合溶液により、in vitroのモデル細胞骨格を構築した。モデル細胞骨格のナノメートルスケールからマイクロメートルスケールでの構造情報を中性子小角散乱と蛍光顕微鏡を併用することにより定量的に調べた。本研究では、細胞内部で塩濃度の揺らぎが存在し、アクチン濃度が高い点を踏まえ、高アクチン濃度条件で塩濃度変化による階層構造の変化を詳細に追跡した。顕微鏡観察から、塩濃度が低い場合には、コイル構造、塩濃度が高くなるとバンドル構造へとその形態を変化させるが、その際に、バンドル内のアクチン間距離やパッキング状態などのナノオーダーの微細構造を変えながら、バンドルの太さを変化させていることを明らかにした。得られた定量的なパラメータから、構造形成要因について詳細に検討した。
増井 友美; 敷中 一洋*; 小泉 智; 角五 彰*; 橋本 竹治; Gong, J.*
no journal, ,
アクチンは細胞運動に重要な役割を果たしているタンパク質であり、アクチン結合タンパク質ともに形成する細胞骨格が細胞運動を担う。本研究では、アクチンとアクチン結合タンパク質間の静電相互作用に着目し、アクチン結合タンパク質の代わりにポリカチオンを用い、塩濃度変化によるアクチン・ポリカチオン複合体の階層構造を、中性子小角散乱法と蛍光顕微鏡法により得た。その結果、アクチン・ポリカチオン複合体は、一定サイズのプロトバンドル,プロトバンドルからなるスーパーバンドルと階層的に凝縮していることを明らかにした。溶液中の塩濃度上昇に伴って、プロトバンドル内のアクチンの配列秩序の向上,プロトバンドルの太さ成長が観測され、同時に、蛍光顕微鏡で観測される高次構造は、グロビュール状からバンドルへと変化した。さらに塩濃度を上げると、複合体は単一アクチンへと溶解した。塩濃度の増大は、アクチン間斥力を緩和し、プロトバンドルの太さ成長を引き起こす。その結果、曲げ弾性エネルギーが増大し、グロビュール状からバンドルへと形態変化する。このようなダイナミックな構造変化は、細胞運動における塩濃度変化の重要性を示唆する。

