


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
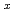 Fe
Fe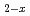 GeO
GeO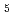 with andalusite structure
with andalusite structure柿本 和勇*; 高田 早紀*; 太田 寛人*; 原口 祐哉*; 萩原 雅人; 鳥居 周輝*; 神山 崇*; 三田村 裕幸*; 徳永 将史*; 畠山 温*; et al.
Journal of the Physical Society of Japan, 91(5), p.054704_1 - 054704_7, 2022/05
被引用回数:1 パーセンタイル:15.48(Physics, Multidisciplinary)The magnetism of AlFeGeO from 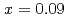 to
to 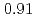 with the andalusite structure was examined. AlFeGeO at low temperatures was found to show a weak ferromagnet-like behavior for
with the andalusite structure was examined. AlFeGeO at low temperatures was found to show a weak ferromagnet-like behavior for 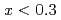 and a spin glass behavior for
and a spin glass behavior for 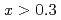 . The small spontaneous magnetization observed in the weak ferromagnet-like samples may be caused by the existence of Dzyaloshinsky-Moriya interaction and the difference in the magnitude of the magnetic moments of Fe
. The small spontaneous magnetization observed in the weak ferromagnet-like samples may be caused by the existence of Dzyaloshinsky-Moriya interaction and the difference in the magnitude of the magnetic moments of Fe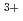 in the octahedral and trigonal bipyramidal sites. The appearance of the spin glass behavior implies that the dilution of Fe ions by Al ions in AlFeGeO causes the competition between ferromagnetic and antiferromagnetic interactions. The
in the octahedral and trigonal bipyramidal sites. The appearance of the spin glass behavior implies that the dilution of Fe ions by Al ions in AlFeGeO causes the competition between ferromagnetic and antiferromagnetic interactions. The 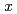 dependence of the site occupancy of Fe ions suggests that Fe
dependence of the site occupancy of Fe ions suggests that Fe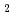 GeO with the andalusite structure cannot be synthesized.
GeO with the andalusite structure cannot be synthesized.
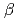 -lactamaseのCs
-lactamaseのCs 選択性結合部位の発見
選択性結合部位の発見新井 栄揮; 柴崎 千枝; 清水 瑠美; 安達 基泰; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
九州シンクロトロン光研究センター年報,2014, p.17 - 19, 2016/03
タンパク質は生物の生命活動のために、金属イオンの電荷数の違いやイオン半径のわずかな違いを識別して結合するなど、緻密な原子・分子認識機構を有する。多くのタンパク質が必須元素の金属を結合することは広く知られている。しかし、タンパク質が必須元素ではないセシウム(Cs)を選択して結合しうるのか、また、その結合部位はどのような構造をしているのか等は明らかにされていなかった。特に、東京電力福島第一原子力発電所事故によって放出された放射性Csの生物への影響を知るためや、タンパク質を利用したCs吸着剤を開発するためには、タンパク質におけるCsの結合のしやすさや結合部位の構造を解明することが重要と考えられた。そこで我々は、好塩性タンパク質とCsの相互作用の研究に着手した。好塩性タンパク質は、塩湖・岩塩・塩蔵食品・発酵食品などの高塩濃度環境に生息する好塩性細菌が作るタンパク質である。好塩性タンパク質は、高塩濃度環境に適応するために多く酸性アミノ酸を含有し、多くの負電荷を有することから、Csを含む様々な金属イオンを結合する可能性があると我々は考えた。本研究では、好塩性タンパク質の中でも比較的高い酸性アミノ酸含量を有し[(Asp + Glu) / (Arg + Lys) = 2.11]、かつ、大腸菌への遺伝子組換えによって大量調製が可能な好塩菌 sp. 560由来-lactamase (以下、HaBLA)について、立体構造の解明とCs結合部位の検出を試みた。
sp. 560由来-lactamase (以下、HaBLA)について、立体構造の解明とCs結合部位の検出を試みた。
 sp. AS-131 isolated from Antarctic Ocean
sp. AS-131 isolated from Antarctic Ocean米澤 悌*; 永山 あい子*; 徳永 廣子*; 石橋 松二郎*; 新井 栄揮; 黒木 良太; 渡邉 啓一*; 荒川 力*; 徳永 正雄*
Protein Journal, 34(4), p.275 - 283, 2015/08
被引用回数:4 パーセンタイル:9.94(Biochemistry & Molecular Biology)好冷菌 sp. AS-131由来ヌクレオシド二リン酸キナーゼ(ASNDK)の大腸菌による発現、および、精製に成功した。通常細菌由来ヌクレオシド二リン酸キナーゼと比較して、ASNDKは、(1)37 Cの大腸菌発現では変性した不溶性の状態で発現する。(2)活性の至適温度は30C低い。など、熱的に不安定な特徴を有していた。また、ASNDKは、中度好塩菌由来ヌクレオシド二リン酸キナーゼのような二量体構造を形成することが示唆された。
Cの大腸菌発現では変性した不溶性の状態で発現する。(2)活性の至適温度は30C低い。など、熱的に不安定な特徴を有していた。また、ASNDKは、中度好塩菌由来ヌクレオシド二リン酸キナーゼのような二量体構造を形成することが示唆された。
-lactamase from the moderate halophile sp.560 and the discovery of a Cs-selective binding site新井 栄揮; 米澤 悌*; 岡崎 伸生*; 松本 富美子*; 柴崎 千枝; 清水 瑠美; 山田 貢*; 安達 基泰; 玉田 太郎; 河本 正秀*; et al.
Acta Crystallographica Section D, 71(3), p.541 - 554, 2015/03
被引用回数:8 パーセンタイル:52.42(Biochemical Research Methods)蛋白質を利用した希少・有害金属捕集材料の研究開発の一環として、中度好塩菌Chromohalobacter sp.560由来・高酸性-Lactamase(HaBLA)のX線結晶構造を解明するとともに、X線異常分散測定により、HaBLA分子上のCs, Sr 結合部位の抽出を試みた。PFのNW3AにてHaBLAのX線結晶構造を解明した後、Cs吸収端(
結合部位の抽出を試みた。PFのNW3AにてHaBLAのX線結晶構造を解明した後、Cs吸収端(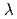 =2.175
=2.175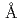 )近傍のX線を利用できるSAGA-LSのBL7やPFのBL17A、及び、Sr吸収端(=0.770)近傍のX線を利用できるSPring-8のBL38B1やPFのBL5Aなどを使用して、HaBLA分子に結合したCs及びSrを同定した。その結果、HaBLA分子上に少なくとも1ヶ所のCs結合部位、3ヶ所のSr結合部位を発見した。特に、今回発見したCs結合部位は、NaがCsの9倍量存在する条件下(Na/Cs = 90mM/10mM)でもCsを選択的に結合できることが明らかになった。このCs選択的結合部位は、Trp側鎖のベンゼン環によるカチオン-
)近傍のX線を利用できるSAGA-LSのBL7やPFのBL17A、及び、Sr吸収端(=0.770)近傍のX線を利用できるSPring-8のBL38B1やPFのBL5Aなどを使用して、HaBLA分子に結合したCs及びSrを同定した。その結果、HaBLA分子上に少なくとも1ヶ所のCs結合部位、3ヶ所のSr結合部位を発見した。特に、今回発見したCs結合部位は、NaがCsの9倍量存在する条件下(Na/Cs = 90mM/10mM)でもCsを選択的に結合できることが明らかになった。このCs選択的結合部位は、Trp側鎖のベンゼン環によるカチオン-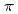 相互作用、および、主鎖の2つの酸素原子によってCsを結合していた。本研究で得たCs結合部位の立体構造情報は、原発事故によって放出された放射性Csを捕集する蛋白質材料の設計(人工的Cs結合部位の設計)の土台として利用できる。
相互作用、および、主鎖の2つの酸素原子によってCsを結合していた。本研究で得たCs結合部位の立体構造情報は、原発事故によって放出された放射性Csを捕集する蛋白質材料の設計(人工的Cs結合部位の設計)の土台として利用できる。
 sp.593
sp.593新井 栄揮; 米澤 悌*; 石橋 松二郎*; 松本 富美子*; 安達 基泰; 玉田 太郎; 徳永 廣子*; Blaber, M.; 徳永 正雄*; 黒木 良太
Acta Crystallographica Section D, 70(3), p.811 - 820, 2014/03
被引用回数:13 パーセンタイル:63.41(Biochemical Research Methods)中度好塩菌 sp.593のペリプラズム蛋白質Alkaline phosphatase(HaAP)は、他の好塩性Alkaline phosphataseと異なり、幅広い塩濃度域(1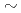 4M NaCl)において機能発現が可能である。そこで本研究では、HaAPの構造学的特徴と好塩性の関係を理解するために、HaAPのX線結晶解析を行った。分解能2.1, 空間群
4M NaCl)において機能発現が可能である。そこで本研究では、HaAPの構造学的特徴と好塩性の関係を理解するために、HaAPのX線結晶解析を行った。分解能2.1, 空間群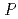 2
2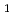 , 格子定数
, 格子定数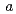 =52.7,
=52.7, 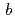 =147.0,
=147.0, 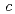 =58.3,
=58.3, 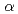 =90, =105.2,
=90, =105.2, 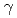 =90, R
=90, R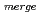 8.4%の回折データを取得して、生物学的構造単位であるHaAP二量体の立体構造を解明することに成功した。また、HaAPの立体構造を、PDB中で最も配列相同性が高い低度好塩菌
8.4%の回折データを取得して、生物学的構造単位であるHaAP二量体の立体構造を解明することに成功した。また、HaAPの立体構造を、PDB中で最も配列相同性が高い低度好塩菌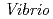 sp.由来VAP(identity 70.0%)の立体構造と比較した。その結果、ASA
sp.由来VAP(identity 70.0%)の立体構造と比較した。その結果、ASA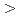 0
0 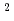 の酸性アミノ酸(D, E)の数は、VAP(57個)よりもHaAP(72個)が多いことが明らかになった。また、VAPとHaAPを構成する疎水性アミノ酸(V, L, I, P, F, M, W)に着目すると、二量体界面に位置する疎水性アミノ酸の数はほぼ同じ(39個と40個)であったが、分子内部(ASAが0)の疎水性アミノ酸はそれぞれ24個と37個であった。このようなHaAPにおける分子表面の高い酸性アミノ酸含量や分子内部の高い疎水性アミノ酸含量は、中度好塩菌のペリプラズム特有の幅広い塩濃度環境下(0.5M飽和塩濃度)における高い可溶性と機能発現の両立に寄与していると考えられる。
の酸性アミノ酸(D, E)の数は、VAP(57個)よりもHaAP(72個)が多いことが明らかになった。また、VAPとHaAPを構成する疎水性アミノ酸(V, L, I, P, F, M, W)に着目すると、二量体界面に位置する疎水性アミノ酸の数はほぼ同じ(39個と40個)であったが、分子内部(ASAが0)の疎水性アミノ酸はそれぞれ24個と37個であった。このようなHaAPにおける分子表面の高い酸性アミノ酸含量や分子内部の高い疎水性アミノ酸含量は、中度好塩菌のペリプラズム特有の幅広い塩濃度環境下(0.5M飽和塩濃度)における高い可溶性と機能発現の両立に寄与していると考えられる。
sp. nucleoside diphosphate kinase新井 栄揮; 米澤 悌; 岡崎 伸生; 松本 富美子; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; Blaber, M.; 徳永 正雄*; 黒木 良太
Protein Science, 21(4), p.498 - 510, 2012/04
被引用回数:15 パーセンタイル:34.81(Biochemistry & Molecular Biology)さまざまな生物種で保存されているヌクレオシドニリン酸キナーゼ(NDK)は、4量体もしくは6量体構造を形成することが知られる。一方、中度好塩菌 sp. 593由来NDK(HaNDK)はNDKとしては例外的に2量体を形成し、E134A変異導入により4量体へ変換される。本研究では、ゲルろ過光散乱及びX線結晶解析により、中度好塩菌 sp. 593由来NDKにおけるE134A変異導入による多量体変換の機構を解明した。また、E134A変異型HaNDKの結晶中には、グラム陰性菌由来MxNDKに類似した4量体構造と大腸菌由来EcNDKに類似した4量体構造が交互に現れることを明らかにした。一般に蛋白質は会合することで熱安定性や基質親和性を増大することから、少ない変異導入による多量体構造の変化は、NDKがさまざまな環境に適合するために有効に寄与している可能性がある。
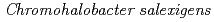 DSM3043; Both residues 134 and 136 are critical for the tetramer assembly
DSM3043; Both residues 134 and 136 are critical for the tetramer assembly徳永 廣子*; 伊豆津 健一*; 新井 栄揮; 米澤 悌; 黒木 良太; 荒川 力*; 徳永 正雄*
Enzyme and Microbial Technology, 46(2), p.129 - 135, 2010/02
被引用回数:6 パーセンタイル:19.99(Biotechnology & Applied Microbiology)本論文では中度好塩菌由来CsNDKの多量体構造にかかわる134及び136番目残基の役割について議論する。CsNDKのGly134やGlu136をAlaやThrに置換した数種類の変異蛋白質を調製し、それらの会合状態を比較した結果、134及び136番目残基の両者がCsNDKのサブユニットの会合に寄与していることが明らかになった。Gly134をAla、Glu136をThrに置換したCsNDK/ANTでは、Ala134が疎水性クラスターを形成することで二量体-二量体の会合が安定化することが判明した。
徳永 廣子*; 石橋 松二郎*; 有坂 文雄*; 新井 栄揮; 黒木 良太; 荒川 力*; 徳永 正雄*
FEBS Letters, 582(7), p.1049 - 1054, 2008/04
被引用回数:17 パーセンタイル:39.33(Biochemistry & Molecular Biology)ヌクレオシド二リン酸化キナーゼ(NDK)は、中度好塩菌由来HaNDKの場合は2量体を形成し、通常細菌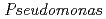 由来PaNDKの場合は4量体を形成する。しかし、HaNDKのGlu134にAlaを変異導入すると4量体を形成するようになり、PaNDKのAla134にGluを変異導入すると2量体を形成するようになった。この結果は、134番目のアミノ酸一残基のみの置換でNDKの複合体構造を変換できることを意味する。中度好塩菌
由来PaNDKの場合は4量体を形成する。しかし、HaNDKのGlu134にAlaを変異導入すると4量体を形成するようになり、PaNDKのAla134にGluを変異導入すると2量体を形成するようになった。この結果は、134番目のアミノ酸一残基のみの置換でNDKの複合体構造を変換できることを意味する。中度好塩菌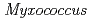 由来NDKの結晶構造に基づいて作製したHaNDKとPaNDKの分子モデルからは、Glu134による反発的相互作用が4量体中の2量体-2量体間の分裂を促すことが判明した。
由来NDKの結晶構造に基づいて作製したHaNDKとPaNDKの分子モデルからは、Glu134による反発的相互作用が4量体中の2量体-2量体間の分裂を促すことが判明した。
新井 栄揮; 米澤 悌*; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
中度好塩菌 sp.593のペリプラズム蛋白質Alkaline phosphatase(HaALP)は、他の好塩性Alkaline phosphataseと異なり、幅広い塩濃度域(14M NaCl)において機能発現が可能である。そこで本研究では、HaALPの構造学的特徴と好塩性の関係を理解するために、HaALPのX線結晶解析を行った。分解能2.1、空間群2、格子定数=52.7, =147.0, =58.3, =90, =105.2, =90, R 8.4%の回折データを取得して、生物学的構造単位であるHaALP二量体の立体構造を解明することに成功した。また、HaALPの立体構造を、PDB中で最も配列相同性が高い低度好塩菌 sp.由来VALP (identity 70.0%)の立体構造と比較した。その結果、ASA0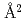 の酸性アミノ酸(D, E)の数は、VALP (57個)よりもHaALP (72個)が多いことが明らかになった。また、VALPとHaALPを構成する疎水性アミノ酸(V, L, I, P, F, M, W)に着目すると、二量体界面に位置する疎水性アミノ酸の数はほぼ同じ(39個と40個)であったが、分子内部(ASAが0)の疎水性アミノ酸はそれぞれ24個と37個であった。このようなHaALPにおける分子表面の高い酸性アミノ酸含量や分子内部の高い疎水性アミノ酸含量は、中度好塩菌のペリプラズム特有の幅広い塩濃度環境下(0.5M飽和塩濃度)における高い可溶性と機能発現の両立に寄与していると考えられる。
の酸性アミノ酸(D, E)の数は、VALP (57個)よりもHaALP (72個)が多いことが明らかになった。また、VALPとHaALPを構成する疎水性アミノ酸(V, L, I, P, F, M, W)に着目すると、二量体界面に位置する疎水性アミノ酸の数はほぼ同じ(39個と40個)であったが、分子内部(ASAが0)の疎水性アミノ酸はそれぞれ24個と37個であった。このようなHaALPにおける分子表面の高い酸性アミノ酸含量や分子内部の高い疎水性アミノ酸含量は、中度好塩菌のペリプラズム特有の幅広い塩濃度環境下(0.5M飽和塩濃度)における高い可溶性と機能発現の両立に寄与していると考えられる。
新井 栄揮; 米澤 悌; 岡崎 伸生; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
ヌクレオシドニリン酸キナーゼ(NDK)は、あらゆる生物に保存された蛋白質であり、オリゴマー構造を形成することが知られている。グラム陰性細菌由来のNDKは基本2量体が2つ集まった4量体,真核生物及び古細菌由来のNDKは基本2量体が3つ集まった6量体などの会合構造をとる。本研究では、中度好塩菌 sp. 593株由来NDK(HaNDK)のE134A変異型の結晶構造を調べ、結晶構造中にグラム陰性菌由来NDKに類似した4量体構造(Type I)と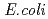 由来NDKに類似した4量体構造(Type II)が交互に現れることを明らかにした。この結果により、HaNDKが少ない変異導入によって多量体構造を容易に変化させることが示唆された。
由来NDKに類似した4量体構造(Type II)が交互に現れることを明らかにした。この結果により、HaNDKが少ない変異導入によって多量体構造を容易に変化させることが示唆された。
新井 栄揮; 米澤 悌*; 安達 基泰; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
好塩性蛋白質は高い酸性アミノ酸含量により分子表面に多くの負電荷を有し、高塩濃度環境下において機能発現や立体構造形成が可能な特殊な蛋白質である。一般に好塩性蛋白質は分子表面の負電荷の影響で結晶化が困難であるが、われわれは二価金属イオンを添加剤とした結晶化スクリーニングを構築し、これまでに数種類の好塩性蛋白質の結晶化及びX線結晶解析に成功している。本会議では、好塩性蛋白質の構造-機能相関の研究の一環として行った sp.560由来-Lactamase(HaBLA)、 sp.593由来nucleoside diphosphate kinase(HaNDK)及びalkaline phosphatase(HaAP)のX線結晶解析の結果について発表する。これら三種類の好塩性蛋白質の立体構造を解明して比較した結果、分子表面における負電荷の密度及び分子内部の疎水性アミノ酸含量はHaAPが最も高いことが判明した。分子表面の負電荷は高塩濃度環境下での可溶性に寄与するとともに、金属イオンとの相互作用の可能性をもたらし、また、分子内部の高い疎水性アミノ酸含量は高塩濃度環境下での蛋白質の構造安定性に寄与することから、HaAPの構造学的特徴は金属イオンを捕集する蛋白質材料を人工創製に利用できる可能性がある。
-lactamaseによるCs及びSr認識機構新井 栄揮; 米澤 悌; 安達 基泰; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
蛋白質は金属イオンの電荷数の違いやイオン半径のわずかな違い(Na 1.14, K 1.52, Mg 0.86, Ca 1.14)などを識別することができる。これまでにわれわれは蛋白質が有する上記の特徴に着目し、蛋白質を利用した希少金属の捕集研究を進めてきたが、原子力発電所事故を背景に、Cs, Sr捕集蛋白質材料の開発に着手した。その一環としてわれわれは、分子表面に数多くの負電荷を有し、多くの金属イオンを結合できる可能性がある sp.560由来・好塩性-Lactamase(HaBLA)について、X線結晶解析によるCs, Sr結合部位の抽出を試みた。0.1M Cs, 0.2M Srを含む結晶化溶液から得られたHaBLA結晶について、空間群3,分解能2.0,格子定数a=b=115.9, c=67.9, Rmerge 9.6%の回折データを取得し、非対称単位(蛋白質3分子)あたり3つのCs結合部位、及び、6つのSr結合部位の同定に成功した。Cs, Sr選択性の高い金属結合部位を抽出できれば、その構造をスカフォールドとしてCs, Sr結合部位の人工創製が可能になると期待できる。
徳永 廣子*; 石橋 松二郎*; 有坂 文雄*; 新井 栄揮; 黒木 良太; 山口 類*; 荒川 力*; 徳永 正雄*
no journal, ,
ヌクレオシド二リン酸化キナーゼ(NDK)は、ほとんどの生物種に広く保存された酵素である。グラム陰性中度好塩菌 sp. 593由来のNDK(HaNDK)とグラム陰性非好塩菌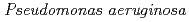 由来NDK(PaNDK)は高い相同性(78% identity, 89% similarity)を示すにもかかわらず、前者は2量体構造、後者は4量体構造を形成する。両者のアミノ酸配列の比較や立体構造予測から、134番目のアミノ酸残基が複合体構造の違いに大きく関与していると予測された。また、HaNDKはGlu134をAlaに置換すると4量体を形成するようになり、逆にPaNDKはAla134をGluに置換すると2量体を形成することが判明した。これらの結果から、野生型HaNDK及び変異型PaNDKは、Glu134のマイナス電荷による反発的相互作用により、4量体形成が阻害されていることが示唆された。
由来NDK(PaNDK)は高い相同性(78% identity, 89% similarity)を示すにもかかわらず、前者は2量体構造、後者は4量体構造を形成する。両者のアミノ酸配列の比較や立体構造予測から、134番目のアミノ酸残基が複合体構造の違いに大きく関与していると予測された。また、HaNDKはGlu134をAlaに置換すると4量体を形成するようになり、逆にPaNDKはAla134をGluに置換すると2量体を形成することが判明した。これらの結果から、野生型HaNDK及び変異型PaNDKは、Glu134のマイナス電荷による反発的相互作用により、4量体形成が阻害されていることが示唆された。
sp.593 (HaNDK)新井 栄揮; 米澤 悌; 岡崎 伸生; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
ヌクレオシドニリン酸キナーゼNDKは、あらゆる生物に保存された蛋白質であり、オリゴマー構造を形成することが知られている。グラム陰性細菌由来のNDKは基本2量体が2つ集まった4量体、真核生物及び古細菌由来のNDKは基本2量体が3つ集まった6量体などの会合構造をとる。本研究では、中度好塩菌由来HaNDKのE134A変異型の結晶構造を調べ、結晶構造中にグラム陰性菌由来MxNDKに類似した4量体構造と大腸菌由来EcNDKに類似した4量体構造が交互に現れることを明らかにした。この結果により、NDKが少ない変異導入によって多量体構造を容易に変化させることが示唆された。
-Lactamase derived from sp.560新井 栄揮; 徳永 廣子*; 玉田 太郎; 米澤 悌; 安達 基泰; 山田 貢; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
好塩性蛋白質は、表面に存在する多くの酸性アミノ酸残基によってさまざまな無機イオンを結合することができる。好塩性蛋白質はレアメタルや放射性金属イオンの捕集材料として用いることができる可能性があるため、分子構造・機能研究グループでは好塩性蛋白質の分子構造を研究している。最近、分子構造・機能研究グループでは、中度好塩菌 sp.560由来-Lactamase(HaBLA)のX線結晶解析に成功した。Photon FactoryのNE3Aビームラインにより、3.0分解能の回折データを収集し、構造解析を行った結果、HaBLAの主鎖構造は非好塩性-Lactamaseと類似した構造をとるが、HaBLAの分子表面は大部分が負電荷で占められることが判明した。このような構造学的情報は、CsやSrのような金属の特異性を向上させるために有効となる。
sp.560株由来-LactamaseのX線結晶解析新井 栄揮; 米澤 悌; 岡崎 伸生; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
近年、希少金属の供給の不安定化に伴って、海水等から希少金属イオンを効率的に回収する技術の確立が求められている。われわれは、分子表面に数多くの負電荷を有する好塩性蛋白質を、希少金属を捕集する材料として着目し、数種類の好塩性蛋白質のX線結晶解析を実施し、無機イオンの結合にかかわる蛋白質分子表面の構造学的特徴(極性原子の配置や配位数など)の解析を行っている。その研究の一環として、われわれは中度好塩菌 sp.560由来・好塩性-LactamaseのX線結晶解析を行い、立体構造決定に成功した。現在、構造解析結果について極性原子の配置や配位数などを精査し、NaやMgなどが結合しうる部位の抽出を行っている。これらの金属結合部位に変異導入による電荷や構造的な摂動を与えて無機イオン選択性を操作できれば、希少金属結合部位を人工的に創製することも可能になると考えられる。この技術が確立されれば、原子力発電所の事故により漏出した放射性セシウム等の回収にも応用できると期待される。
-lactamaseのCs選択性結合部位の発見新井 栄揮; 安達 基泰; 河本 正秀*; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
蛋白質を利用した希少金属・有害金属捕集材料の研究開発の一環として、X線異常分散測定により、 sp.560由来・好塩性-Lactamase(HaBLA)の結晶構造中のCs, Sr結合部位の抽出を試みた。Csの同定にはCs吸収端(=2.175)近傍のX線を利用できるSAGA-LSのBL7、Srの同定にはSr吸収端(=0.770)近傍のX線を利用できるSPring-8のBL38B1およびPFのNW12Aを使用した。本測定・解析の結果、HaBLAの結晶構造中にCsやSrに由来する異常分散差フーリエ電子密度を観測し、HaBLA分子上に少なくとも1ヶ所のCs結合部位、3ヶ所のSr結合部位を発見した。特に、今回発見したCs結合部位は、NaがCsの9倍量存在する条件下(Na/Cs=90mM/10mM)でもCsを選択的に結合できることが明らかになった。このCs選択的結合部位は、Trp側鎖のベンゼン環によるカチオン-相互作用、および、主鎖の2つの酸素原子によってCsを結合していた。本研究で得たCs, Sr結合部位の立体構造情報は、福島第一原子力発電所事故によって放出された放射性Cs, Srを捕集する蛋白質材料の設計(人工的Cs, Sr結合部位の設計)の土台として利用できる。
-ラクタマーゼの立体構造と低塩濃度における酵素活性維持との相関新井 栄揮; 安達 基泰; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
一般的に、豊富な酸性アミノ酸含量を有する好塩性蛋白質は、低塩濃度下では構造が不安定化し、酵素活性などの機能を失う。しかし、中度好塩菌の生息域では、ペリプラズムは0.5M飽和塩濃度の環境にあり、そこに局在する好塩性蛋白質は低塩濃度への適合も要求される。今回我々は、中度好塩菌ペリプラズム酵素の構造・機能研究の一環として、中度好塩菌 sp.560由来ペリプラズム酵素HaBLAの立体構造をX線結晶回折法によって解析するとともに、その酵素活性を等温滴定熱量測定法により解析した。HaBLAは04M NaClにおいてペニシリンGを効果的に分解した。その分解活性は、2M NaCl存在下ではkcat/Km=142.8(S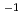
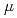 M )であり、14M NaClの範囲でその50%を維持し、0M NaClでも20%を維持していた。そこで我々は、HaBLA結晶構造を精査し、機能発現に関わる構造的特徴を調べた。その結果、HaBLA表面は大部分が負電荷で占められるが、活性部位周辺に局所的に正電荷を有するとともに、活性部位入口に疎水性クラスターを有することを発見した。ペニシリンGは負電荷を有するともに芳香環も有する。HaBLAは活性部位周辺の正電荷による静電相互作用と疎水性クラスターによる疎水性相互作用によってペニシリンGを活性部位へ誘導し、低塩濃度での活性を維持していると考えられる。
M )であり、14M NaClの範囲でその50%を維持し、0M NaClでも20%を維持していた。そこで我々は、HaBLA結晶構造を精査し、機能発現に関わる構造的特徴を調べた。その結果、HaBLA表面は大部分が負電荷で占められるが、活性部位周辺に局所的に正電荷を有するとともに、活性部位入口に疎水性クラスターを有することを発見した。ペニシリンGは負電荷を有するともに芳香環も有する。HaBLAは活性部位周辺の正電荷による静電相互作用と疎水性クラスターによる疎水性相互作用によってペニシリンGを活性部位へ誘導し、低塩濃度での活性を維持していると考えられる。
選択性結合部位の発見新井 栄揮; 安達 基泰; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
数多くの金属イオンと相互作用する可能性がある好塩性タンパク質分子を利用して、希少金属・有害金属イオンの結合部位を見出す研究を進めてきた。今回我々は、X線結晶解析により、中度好塩菌 sp.560由来・HaBLAの立体構造を解明するとともに、X線異常分散測定により、HaBLA結晶構造中のCs, Sr結合部位の抽出を試みた。Csの同定にはSAGA-LSのBL7、Sr+の同定にはSPring-8のBL38B1およびPFのNW12Aを使用した。本研究の結果、HaBLAの結晶構造中に、Csに由来する17.5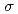 レベル以上、Srに由来する7.0レベル以上の異常分散差フーリエ電子密度を見出し、HaBLA分子上に少なくとも1ヶ所のCs結合部位、3ヶ所のSr結合部位を発見した。特に、今回発見したCs結合部位は、Trp側鎖のベンゼン環によるカチオン-相互作用、および、主鎖の2つの酸素原子によってCsを結合しており、NaがCsの9倍量存在する条件下(Na/Cs = 90mM/10mM)でもCsを選択的に結合できることが明らかになった。
レベル以上、Srに由来する7.0レベル以上の異常分散差フーリエ電子密度を見出し、HaBLA分子上に少なくとも1ヶ所のCs結合部位、3ヶ所のSr結合部位を発見した。特に、今回発見したCs結合部位は、Trp側鎖のベンゼン環によるカチオン-相互作用、および、主鎖の2つの酸素原子によってCsを結合しており、NaがCsの9倍量存在する条件下(Na/Cs = 90mM/10mM)でもCsを選択的に結合できることが明らかになった。
新井 栄揮; 米澤 悌; 岡崎 伸生; 玉田 太郎; 徳永 廣子*; 石橋 松二郎*; 徳永 正雄*; 黒木 良太
no journal, ,
ヌクレオシドニリン酸キナーゼ(NDK)は、あらゆる生物に保存された蛋白質であり、オリゴマー構造を形成することが知られている。グラム陰性細菌由来のNDKは基本2量体が2つ集まった4量体、真核生物及び古細菌由来のNDKは基本2量体が3つ集まった6量体などの会合構造をとる。本研究では、中度好塩菌 sp. 593株由来NDK(HaNDK)のE134A変異型の結晶構造を調べ、結晶構造中にグラム陰性菌由来NDKに類似した4量体構造(Type I)と由来NDKに類似した4量体構造(Type II)が交互に現れることを明らかにした。この結果により、HaNDKが少ない変異導入によって多量体構造を容易に変化させることが示唆された。
